Takes of Marine Mammals Incidental to Specified Activities; Taking Marine Mammals Incidental to a Marine Geophysical Survey Off Western Mexico in the Eastern Tropical Pacific Ocean
NMFS has received a request from the Lamont-Doherty Earth Observatory (L-DEO) for authorization to take marine mammals incidental to a marine geophysical survey off Western Mexi...
National Marine Fisheries Service (NMFS), National Oceanic and Atmospheric Administration (NOAA), Commerce.
ACTION:
Notice; proposed incidental harassment authorization; request for comments on proposed authorization and possible renewal.
SUMMARY:
NMFS has received a request from the Lamont-Doherty Earth Observatory (L-DEO) for authorization to take marine mammals incidental to a marine geophysical survey off Western Mexico in the Eastern Tropical Pacific Ocean (ETP). Pursuant to the Marine Mammal Protection Act (MMPA), NMFS is requesting comments on its proposal to issue an incidental harassment authorization (IHA) to incidentally take marine mammals during the specified activities. NMFS is also requesting comments on a possible one-time, 1-year renewal that could be issued under certain circumstances and if all requirements are met, as described in Request for Public Comments at the end of this notice. NMFS will consider public comments prior to making any final decision on the issuance of the requested MMPA authorization and agency responses will be summarized in the final notice of our decision.
DATES:
Comments and information must be received no later than June 4, 2025.
Instructions:
NMFS is not responsible for comments sent by any other method, to any other address or individual, or received after the end of the comment period. Comments, including all attachments, must not exceed a 25-megabyte file size. All comments received are a part of the public record and will generally be posted online at
https://www.fisheries.noaa.gov/permit/incidental-take-authorizations-under-marine-mammal-protection-act
without change. All personal identifying information (
e.g.,
name, address) voluntarily submitted by the commenter may be publicly accessible. Do not submit confidential business information or otherwise sensitive or protected information.
FOR FURTHER INFORMATION CONTACT:
Jenna Harlacher, Office of Protected Resources, NMFS, (301) 427-8401.
SUPPLEMENTARY INFORMATION:
Background
The MMPA prohibits the “take” of marine mammals, with certain exceptions. Sections 101(a)(5)(A) and (D) of the MMPA (16 U.S.C. 1361et seq.) direct the Secretary of Commerce (as delegated to NMFS) to allow, upon request, the incidental, but not intentional, taking of small numbers of marine mammals by U.S. citizens who engage in a specified activity (other than commercial fishing) within a specified geographical region if certain findings are made and either regulations are proposed or, if the taking is limited to harassment, a notice of a proposed IHA is provided to the public for review.
Authorization for incidental takings shall be granted if NMFS finds that the taking will have a negligible impact on the species or stock(s) and will not have an unmitigable adverse impact on the availability of the species or stock(s) for taking for subsistence uses (where relevant). Further, NMFS must prescribe the permissible methods of taking and other “means of effecting the least practicable adverse impact” on the affected species or stocks and their habitat, paying particular attention to rookeries, mating grounds, and areas of similar significance, and on the availability of the species or stocks for taking for certain subsistence uses (referred to in shorthand as “mitigation”); and requirements pertaining to the monitoring and reporting of the takings. The definitions of all applicable MMPA statutory terms used above are included in the relevant sections below and can be found in section 3 of the MMPA (16 U.S.C. 1362) and NMFS regulations at 50 CFR 216.103.
National Environmental Policy Act
To comply with the National Environmental Policy Act of 1969 (NEPA; 42 U.S.C. 4321et seq.) and NOAA Administrative Order (NAO) 216-6A, NMFS must review our proposed action (
i.e.,
the issuance of an IHA) with respect to potential impacts on the human environment.
This action is consistent with categories of activities identified in Categorical Exclusion B4 (IHAs with no anticipated serious injury or mortality) of the Companion Manual for NAO 216-6A, which do not individually or cumulatively have the potential for significant impacts on the quality of the human environment and for which we have not identified any extraordinary circumstances that would preclude this categorical exclusion. Accordingly, NMFS has preliminarily determined that the issuance of the proposed IHA qualifies to be categorically excluded from further NEPA review.
Summary of Request
On November 14, 2024, NMFS received a request from the L-DEO for an IHA to take marine mammals incidental to a marine geophysical survey off Western Mexico in the ETP. After sending initial questions to the applicant, L-DEO deferred the IHA request due to vessel clearance issues. On January 29, 2025 L-DEO alerted us that they received clearance but their proposed survey dates had changed. With revised dates, NMFS continued processing the application and it was deemed adequate and complete on March 4, 2025. L-DEO's request is for take of 29 species of marine mammals, by Level B harassment only. Neither L-DEO nor NMFS expect serious injury or mortality to result from this activity and, therefore, an IHA is appropriate.
Description of Proposed Activity
Overview
Researchers from the New Mexico Institute of Mining and Technology (NMT) and University of New Mexico (UNM), with funding from the National Science Foundation (NSF), and in collaboration with Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), propose to conduct a low-energy marine seismic survey, using airguns as the acoustic source, and heat probe measurements, conducted from the research vessel (R/V)
Marcus G. Langseth
(
Langseth), which is owned and operated by L-DEO. The proposed survey would occur off Western Mexico in the ETP from approximately November 2025 to December 2025. The proposed survey would occur within the Mexican exclusive economic zone (EEZ) but outside of territorial waters, in
( printed page 19091)
water depths ranging from approximately 1,000 to 5,300 meters (m). To complete this 2-dimensional (2-D) survey, the
Langseth
would tow a cluster of two 45 cubic inch (in3
, 737 cubic centimeters (cc)) generator injector (GI) airguns with a total discharge volume of 90 in3
(~1,475 cc) at a depth of 3 m. The airgun array receiver would consist of a 3-5 kilometer (km) long solid-state hydrophone streamer. The airguns would fire at a shot interval of 6.25-12.5 m. Approximately 1,258 kilometers (km) of seismic acquisition is planned. Airgun arrays would introduce underwater sound that may result in take of marine mammals.
The purpose of the proposed survey is to obtain information on the sediment distribution and geologic structure of the Cocos plate and margin wedge, which is necessary for constraining the thermal structure of the subduction zone offshore southern Mexico. The main goal of the proposed seismic surveys is to acquire 2-D seismic reflection data, in conjunction with densely spaced heat probe measurements, to quantify the effects of fluid circulation in oceanic crust on temperatures in the southern Mexico subduction zone.
Dates and Duration
The
Langseth
is proposed to leave out of port in Manzanilla, Mexico, on November 18, 2025, and return to port in Manzanilla, Mexico, on December 15, 2025, after the survey is completed. The survey is expected to last 24 days, which includes approximately 7 days of seismic operations, 14 days of heat probe measurements, and 3 days of transit.
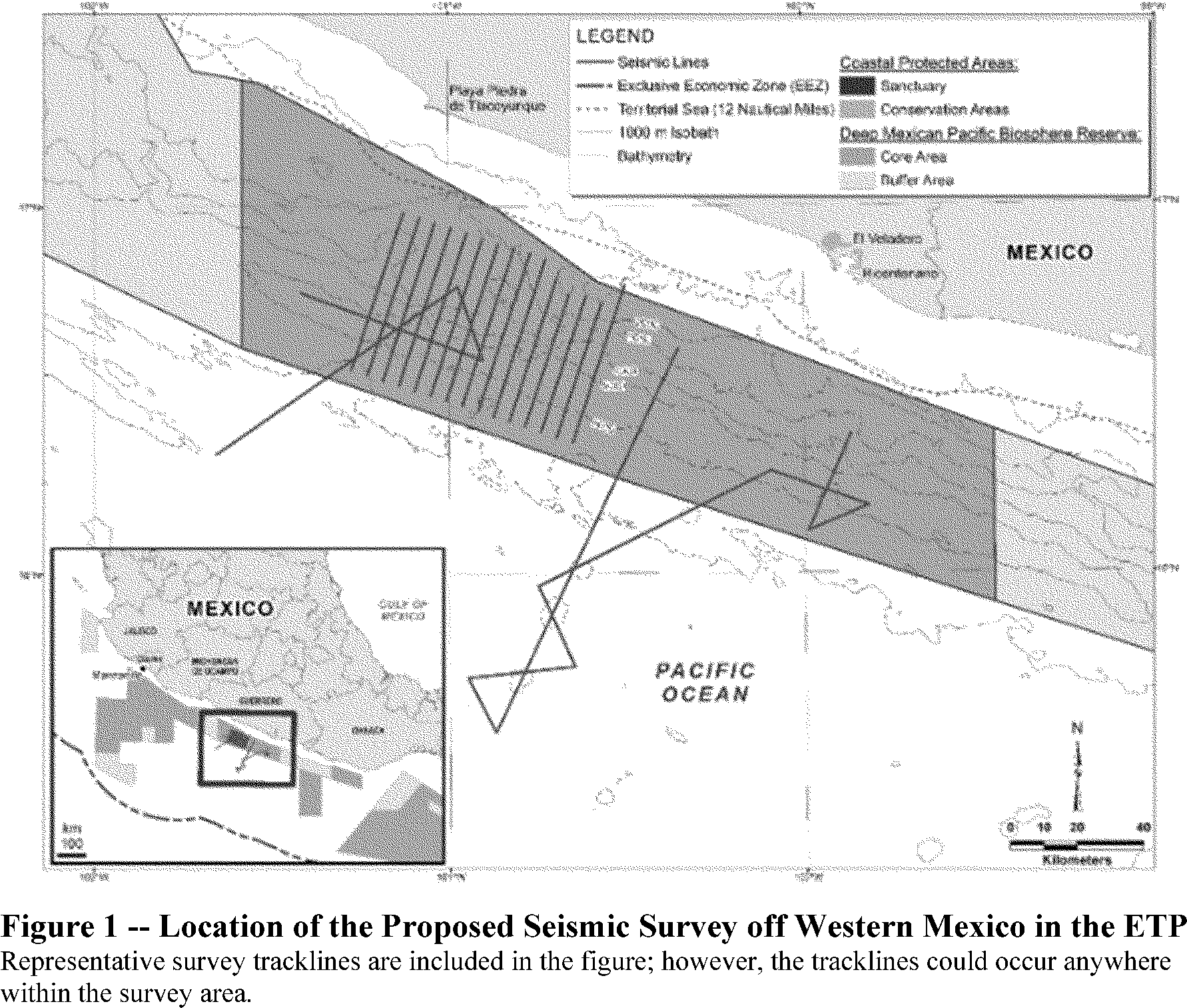
Specific Geographic Region
The proposed survey would occur within approximately ~15.5-17° N and 99.5-102° W, off the Pacific coast of Mexico within the EEZ of Mexico, in water depths ranging from approximately 1,000 to 5,300 m. The region where the survey is proposed to occur is depicted in figure 1. Representative survey tracklines are shown; however, some deviation in actual tracklines, including the order of survey operations, could be necessary for reasons such as science drivers, poor data quality, inclement weather, or mechanical issues with the research vessel and/or equipment. Therefore, for the proposed survey, the tracklines could occur anywhere within the coordinates noted above. The
Langseth
would likely leave out of and return to port in Manzanilla, Mexico (approximately 420 km north of the survey area).
( printed page 19092)
Detailed Description of the Specified Activity
The procedures to be used for the proposed survey would be similar to those used during previous seismic surveys conducted by L-DEO and would use conventional seismic methodology. The survey would involve one source vessel,
Langseth,
which is owned and operated by L-DEO. During the low-energy 2D seismic survey,
Langseth
would tow two GI airguns with a total discharge volume of 90 in3
. The two inline airguns would be spaced 2.46 m apart. The airgun array configurations are illustrated in figure 2-14 of NSF and the U.S. Geological Survey's (USGS) Programmatic Environmental Impact Statement (PEIS; NSF-USGS, 2011). (The PEIS is available online at:
https://www.nsf.gov/geo/oce/envcomp/usgs-nsf-marine-seismic-research/nsf-usgs-final-eis-oeis_3june2011.pdf). The receiving system would consist of a 3-5 km long solid-state hydrophone streamer. As the airgun arrays are towed along the survey lines, the hydrophone streamer would transfer the data to the on-board processing system. Approximately 1,258 km of seismic acquisition are planned. The survey would take place in water depths ranging from approximately 1,000 to 5,300 m.
Heat flow data would be acquired with a heat flow probe that takes the temperature of the sediments like a thermometer. Heat probe measurements are made by lowering the probe through the water column and letting it plunge ~3.5 m into the sediment. Measurements consist of two parts—thermal gradient and conductivity—and would be made every ~500-1,000 m. At each measurement site the probe is left in the seafloor for ~15 minutes. After the measurement is taken, the probe is pulled out of the sediment and raised ~200 m above the seafloor, the ship then moves position along the transect, and the process is repeated (referred to as “pogo” mode). During heat flow probe operations, a 12-kilohertz (kHz) bottom-finding pinger would be employed, and an acoustic release would be used once during an initial calibration of the heat probe activities.
The heat flow probe would be equipped with an ultra-short baseline (USBL) transducer acoustic positioning system (or pinger) to allow it to “talk” with the research vessel. The pole-mounted USBL transducer pings once per second to the receiver to locate the heat flow probe location and vice versa. The reflected pings are picked up by a Knudsen Chirp 3260 sub-bottom profiler (SBP). While on station for heat flow measurements, the MBES would be turned off.
In addition to the operations of the airgun array and the heat flow probe, the ocean floor would be mapped with the Kongsberg EM 122 multibeam echosounder (MBES), and a SBP. A Teledyne RDI 75 kHz Ocean Surveyor acoustic doppler current profiler (ADCP) would be used to measure water current velocities. Take of marine mammals is not expected to occur incidental to use of the MBES, SBP, ADCP, and heat flow probe operations whether or not the airguns are operating simultaneously with the other sources. Given their characteristics (
e.g.,
narrow downward-directed beam), marine mammals would experience no more than one or two brief ping exposures, if any exposure were to occur. NMFS does not expect that the use of these sources is likely to cause take of marine mammals.
Proposed mitigation, monitoring, and reporting measures are described in detail later in this document (please see Proposed Mitigation and Proposed Monitoring and Reporting).
Description of Marine Mammals in the Area of Specified Activities
Sections 3 and 4 of the application summarize available information regarding status and trends, distribution and habitat preferences, and behavior and life history of the potentially affected species. NMFS fully considered all of this information, and we refer the reader to these descriptions, instead of reprinting the information. Additional information about these species (
e.g.,
physical and behavioral descriptions) may be found on NMFS' website (
https://www.fisheries.noaa.gov/find-species). NMFS refers the reader to the aforementioned source for general information regarding the species listed in table 1.
Table 1 lists all species or stocks for which take is expected and proposed to be authorized for this activity and summarizes information related to the population or stock, including regulatory status under the MMPA and Endangered Species Act (ESA) and potential biological removal (PBR), where known. PBR is defined by the MMPA as the maximum number of animals, not including natural mortalities, that may be removed from a marine mammal stock while allowing that stock to reach or maintain its optimum sustainable population (as described in NMFS' stock assessment reports (SARs)). While no serious injury or mortality is anticipated or proposed to be authorized here, PBR and annual serious injury and mortality (M/SI) from anthropogenic sources are included here as gross indicators of the status of the species or stocks and other threats.
Marine mammal abundance estimates presented in this document represent the total number of individuals that make up a given stock or the total number estimated within a particular study or survey area. NMFS's stock abundance estimates for most species represent the total estimate of individuals within the geographic area, if known, that comprises that stock. For some species, this geographic area may extend beyond U.S. waters. All managed stocks in this region are assessed in NMFS's U.S. Pacific SARs. All values presented in table 1 are the most recent available at the time of publication and are available in the draft 2024 SARs (available online at:
https://www.fisheries.noaa.gov/national/marine-mammal-protection/draft-marine-mammal-stock-assessment-reports). Where available, abundance and status information is also presented for marine mammals in the Pacific waters of Mexico and/or the greater ETP region. Table 1 denotes the status of species and stocks under the U.S. MMPA and ESA. We note also that the Guadalupe fur seal is classified as “En peligro de extinción” (in danger of extinction) under the Norma Oficial Mexicana NOM-059-SEMARNAT-2010 and all other marine mammal species listed in table 1, with the exception of Longman's beaked whales and Deraniyagala's beaked whales, are listed as “Sujetas a protección especial” (subject to special protection).
( printed page 19093)
Table 1—Species Likely Impacted by the Specified Activities
Common name
Scientific name
Stock
ESA/MMPA
status;
strategic
(Y/N) 1
Stock
abundance
(CV, N
min
,
most recent
abundance
survey) 2
PBR
Annual
M/SI 3
ETP
abundance 4
Mexico
Pacific
abundance 5
Order Artiodactyla—Cetacea—Mysticeti (baleen whales)
Family Balaenopteridae (rorquals):
Humpback Whale
Megaptera novaeangliae
Central America/Southern Mexico-California-Oregon-Washington
E, D, Y
1,496 (0.2, 1,284, 2021)
3.5
14.9
2,566
Minke whale
Balaenoptera acutorostrata
N/A
-, -, N
N/A
N/A
N/A
115
Bryde's whale
Balaenoptera edeni
Eastern Tropical Pacific
-, -, N
Unknown (Unknown, Unknown, N/A)
Undetermined
Unknown
10,411
649
Sei whale
Balaenoptera borealis
Eastern N Pacific
E, D, Y
519 (0.4, 374, 2014)
0.75
≥0.2
0
Fin whale
Balaenoptera physalus
N/A
E, D, Y
N/A
N/A
N/A
574
145
Odontoceti (toothed whales, dolphins, and porpoises)
Family Physeteridae:
Sperm whale
Physeter macrocephalus
N/A
E, D, Y
N/A
N/A
N/A
4,145
2,810
Family Kogiidae:
Dwarf Sperm Whale
Kogia sima
N/A
N/A
N/A
N/A
N/A
6
11,200
Pygmy Sperm Whale
Kogia breviceps
N/A
-, -, N
N/A
N/A
N/A
6
11,200
Family Ziphiidae (beaked whales):
Cuvier's Beaked Whale
Ziphius cavirostris
N/A
-, -, N
N/A
N/A
N/A
7
20,000
8
68,828
Longman's beaked whale
Indopacetus pacificus
N/A
-, -, N
N/A
N/A
N/A
1,007
Blainville's beaked whale
Mesoplodon densirostris
N/A
-, -, N
N/A
N/A
N/A
9
25,300
8
68,828
Ginkgo-toothed beaked whale
M. ginkgodens
N/A
-, -, N
N/A
N/A
N/A
9
25,300
8
68,828
Deraniyagala's beaked whale
M. hotaula
N/A
-, -, N
N/A
N/A
N/A
9
25,300
8
68,828
Pygmy beaked whale
M. peruvianus
N/A
-, -, N
N/A
N/A
N/A
9
25,300
8
68,828
Family Delphinidae:
Risso's dolphin
Grampus griseus
N/A
-, -, N
N/A
N/A
N/A
110,457
24,084
Rough-toothed dolphin
Steno bredanensis
N/A
-, -, N
N/A
N/A
N/A
107,663
37,511
Common bottlenose dolphin
Tursiops truncatus
N/A
-, -, N
N/A
N/A
N/A
335,834
61,536
Pantropical spotted dolphin
Stenella attenuata
N/A
-, D, N
N/A
N/A
N/A
10
1,297,091
146,296
Spinner dolphin
Stenella longirostris
N/A
-, D, N
N/A
N/A
N/A
10
2,075,871
186,906
Striped dolphin
Stenella coeruleoalba
N/A
-, -, N
N/A
N/A
N/A
964,362
128,867
Short-beaked common dolphin
Delphinus delphis
N/A
-, -, N
N/A
N/A
N/A
3,127,203
283,196
Fraser's dolphin
Lagenodelphis hosei
N/A
-, -, N
N/A
N/A
N/A
7
289,300
Short-finned pilot whale
Globicephala macrorhynchus
N/A
-, -, N
N/A
N/A
N/A
11
589,315
3,348
Killer whale
Orcinus orca
N/A
-, -, N
N/A
N/A
N/A
7
8,500
852
False killer whale
Pseudorca crassidens
N/A
-, -, N
N/A
N/A
N/A
7
39,800
Pygmy killer whale
Feresa attenuata
N/A
-, -, N
N/A
N/A
N/A
7
38,900
Melon-headed whale
Peponocephala electra
N/A
-, -, N
N/A
N/A
N/A
7
45,400
Order Carnivora—Pinnipedia
Family Otariidae (eared seals and sea lions):
Guadalupe fur seal
Arctocephalus townsendi
Mexico
T, D, Y
63,850 (N/A, 57,199, 2013)
1,959
≥10.0
( printed page 19094)
California sea lion
Zalophus californianus
U.S.
-, -, N
257,606 (N/A, 233,515, 2014)
14,011
>321
105,000
1
ESA status: Endangered (E), Threatened (T)/MMPA status: Depleted (D). A dash (-) indicates that the species is not listed under the ESA or designated as depleted under the MMPA. Under the MMPA, a strategic stock is one for which the level of direct human-caused mortality exceeds PBR or which is determined to be declining and likely to be listed under the ESA within the foreseeable future. Any species or stock listed under the ESA is automatically designated under the MMPA as depleted and as a strategic stock.
3
These values, found in NMFS's SARs, represent annual levels of human-caused mortality plus serious injury from all sources combined (
e.g.,
commercial fisheries, ship strike). Annual M/SI often cannot be determined precisely and is in some cases presented as a minimum value or range. A CV associated with estimated mortality due to commercial fisheries is presented in some cases.
4
From NMFS (2015b) unless otherwise noted.
5
Pacific Mexico excluding the Gulf of California (from Gerrodette and Palacios (1996) unless otherwise noted).
6
Estimate for ETP is mostly for
K. sima
but may also include some
K. breviceps
(Wade and Gerrodette 1993).
7
Wade and Gerrodette 1993.
8
Abundance for all ziphiids.
9
This estimate for the ETP includes all species of the genus
Mesoplodon.
10
Includes abundance of several stocks added together.
11
Based on surveys in 2000 (Gerrodette and Forcada 2002).
As indicated above, all 29 species in table 1 temporally and spatially co-occur with the activity to the degree that take is reasonably likely to occur. As the planned survey lines are outside of the U.S. EEZ, they do not directly overlap with the defined ranges for most U.S. managed stocks (Carretta
et al.,
2024). For some species (
e.g.,
Bryde's whale, Guadalupe fur seal; see table 1), animals encountered during the surveys could be from a defined stock under the MMPA but most marine mammals in the survey area do not belong to any defined stock.
Some species could potentially occur in the proposed survey area but are not likely to be encountered due to the rarity of their occurrence. These species include the North Pacific right whale (
Eubalaena japonica), blue whale (
Balaenoptera musculus), gray whale (
Eschrichtius robustus), Hubbs' beaked whale (
Mesoplodon carlhubbsi), Stejneger's beaked whale (
M. stejnegeri), Perrin's beaked whale (
M. perrini), Baird's beaked whale (
Berardius bairdii), vaquita (
Phocoena sinus), harbor porpoise (
Phocoena phocoena), Dall's porpoise (
Phocoenoides dalli), Pacific white-sided dolphin (
Lagenorhynchus obliquidens), and northern right whale dolphin (
Lissodelphis borealis), which all generally occur well outside or north of the proposed survey area (
e.g,
north of the Baja peninsula). Five additional pinniped species are known to occur in the ETP but are considered extralimital in the proposed survey area: The Galápagos sea lion (
Zalophus wollebaeki), Galápagos fur seal (
Arctocephalus galapagoensis), South American fur seal (
A. australis), and the South American sea lion (
Otaria flavescens), which all occur south of the survey area, and the northern elephant seal (
Mirounga angustirostris) which is found north of the survey area.
In addition to what is included in sections 3 and 4 of the IHA application, and NMFS' website, further detail informing the regional occurrence for select species of particular or unique vulnerability (
i.e.,
information regarding ESA listed or MMPA depleted species) is provided below.
Humpback Whale
The Central America distinct population segment (DPS) equates to the Central America/Southern Mexico-CA/OR/WA stock designated under the MMPA and shown in table 1. The endangered Central America DPS may occur within the proposed survey area, based on the timing of the proposed survey (November-December). Humpbacks from the Central America DPS could be migrating through the survey area at the time of the proposed survey.
Whales in the Central America/Southern Mexico-CA/OR/WA stock winter off the coasts of Nicaragua, Honduras, El Salvador, Guatemala, Panama, Costa Rica, and southern Mexico including the states of Oaxaca and Guerrero, with some animals ranging even farther north (Taylor et al. 2021); they summer off California, Oregon, and Washington (Calambokidis et al. 2000).
Nine sightings were made during surveys off the Pacific coast of Mexico in November 2019 (Oedekoven et al. 2021). The central coast of Oaxaca is thought to be a migratory corridor during winter, with whales typically migrating up to 4 km from shore (Heckel et al. 2020). In 2012, 45 sightings were made off Oaxaca (Castillejos-Moguel and Villegas-Zurita 2014
in
Heckel et al. 2020) including feeding behavior (Villegas-Zurita and Castillejos-Moguel 2013
in
Heckel et al. 2020). Feeding has also been observed in Banderas Bay, which is known to be an aggregation area for humpbacks during the winter months (Frish-Jordán et al. 2019). One sighting was made during an L-DEO survey off Guerrero and Michoacán in May-June 2022 (RPS 2022). Although sightings are regularly made within the region during winter, sightings during the proposed fall survey in deep offshore waters are likely to be less common.
Although there are other stocks of humpback whales found in Mexico near the project area, it's likely that humpbacks from the Mexico DPS (Mexico-North Pacific Stock and Mainland Mexico-CA/OR/WA Stock) would still be migrating south from their northern feeding grounds off of the U.S. west coast and are not likely to be found in the survey area. Additionally, the Mexico DPS's winter breeding grounds are north of the proposed survey area in the Revillagigedos Islands. Therefore, we assume that all humpback whales taken by the proposed survey activities would be from the Central America/Southern Mexico-CA/OR/WA stock.
Sei Whale
Sei whales are less common in the survey area but there have been some reports as summarized below. Sei whales are known to occasionally occur in the Gulf of California (Urbán et al. 2014
in
Heckel et al. 2020), as well as off the west coast of the Baja California Peninsula (Heckel et al. 2020). One sighting has been reported for waters off
( printed page 19095)
Nayarit (Urbán et al. 1997, Guerrero et al. 2006
in
Heckel et al. 2020), and another sighting was made near the northern part of the proposed survey area, off Jalisco (Heckel et al. 2020). González et al. (2008) also reported the presence of sei whales off west coast of Mexico south of 23° N. However, neither Ferguson and Barlow (2001) nor Jackson et al. (2004) positively identified any sei whales in Mexican waters during surveys conducted during July-December. RPS (2022) reported two sightings of single sei whales during an L-DEO survey off Guerrero and Michoacán in May-June 2022.
Fin Whale
Fin whale calls are recorded in the North Pacific year-round (
e.g.,
Moore et al. 2006; Stafford et al. 2007, 2009; Edwards et al. 2015). However, fin whales are considered rare in the proposed survey area. No sightings were made in the proposed survey area during July-December surveys during 1986-1996, 2003, or 2019 (Ferguson and Barlow 2001; Jackson et al. 2004; Oedekoven et al. 2021). Similarly, Edwards et al. (2015) reported no sightings or acoustic detections for the proposed survey area, although sightings have been reported for the Gulf of California and a few sightings exist for offshore waters far west of Mexico. However, González et al. (2008) reported the presence of this species off west coast of Mexico south of 23° N, and a sighting has been reported for Banderas Bay (Arroyo 2017). RPS (2022) reported one fin whale sighting during an L-DEO survey off Guerrero and Michoacán in May-June 2022.
Sperm Whale
During summer and fall, sperm whales are widely distributed in the ETP, although they are generally more abundant in deeper “nearshore” waters than far offshore (
e.g.,
Polacheck 1987; Wade and Gerrodette 1993). More than 180 sightings have been reported for the ETP, with the highest concentrations at 10° N-10° S, 80°-100° W (Guerrero et al. 2006). Sightings for Pacific Mexico include records off the Baja California Peninsula and in the Gulf of California (Guerrero et al. 2006; Heckel et al. 2020). During 25,356 km of surveys (excluding the Gulf of California) within the EEZ of Pacific Mexico, during July-December 1986-1990, 1992 and 1993, 46 sightings of sperm whales were made (Gerrodette and Palacios 1996). No sightings were made along the mainland coast of Mexico during July-December surveys in 2003, although one sighting was made off the west coast of Baja California Sur (Jackson et al. 2004). Records also exist for Banderas Bay (Arroyo 2017) and Oaxaca (Pérez and Gordillo 2002
in
Heckel et al. 2020).
Pantropical Spotted Dolphin
The pantropical spotted dolphin is one of the most abundant cetaceans and is distributed worldwide in tropical and some subtropical waters, between ~40° N and 40° S (Jefferson
et al.,
2015). In the ETP, this species ranges from 25° N off the Baja California Peninsula to 17° S, off southern Peru (Perrin and Hohn, 1994). There are two forms of pantropical spotted dolphin (Perrin 2018a): Coastal (
Stenella attenuata graffmani) and offshore (
S. a. attenuata), both of which could occur within the proposed survey area. Along the coast of Latin America, the coastal form typically occurs within 20 km from shore (Urbán 2008
in
Heckel
et al.,
2020). There are currently three recognized stocks of spotted dolphins in the ETP: The coastal stock and two offshore stocks—the northeast and the west/south stocks (Wade and Gerrodette 1993; Leslie
et al.,
2019). Much of what is known about the pantropical spotted dolphin in the ETP is related to the historical tuna purse-seine fishery in that area (Perrin and Hohn 1994). There was an overall stock decline of spotted dolphins from 1960-1980 because of the fishery (Allen 1985). In 1979, the population size of spotted dolphins in the ETP was estimated at 2.9-3.3 million (Allen 1985). For 1986-1990, Wade and Gerrodette (1993) reported an estimate of 2.1 million. Gerrodette and Forcada (2005) noted that the population of offshore northeastern spotted dolphins had not yet recovered from the earlier population declines; possible reasons for the lack of growth were attributed to unreported bycatch, effects of fishing activity on survival and reproduction, and long-term changes in the ecosystem. The abundance estimate for 2006 was ~857,884 northeastern offshore spotted dolphins, and 439,208 western-southern offshore spotted dolphins; the coastal subspecies was estimated at 278,155 and was less affected by fishing activities (Gerrodette
et al.,
2008). In 2004, the mortality rate in the tuna fishery was estimated at 0.03 percent (Bayliff 2004). Perrin (2018a) noted that for the last few years, hundreds of spotted dolphins have been taken in the fishery. Currently, there are ~640,000 northeastern offshore spotted dolphins inhabiting the ETP (Perrin 2018a). This stock is designated as depleted under the MMPA and may be slow to recover due to continued chase and encirclement by the tuna fishery, which may in turn affect reproductive rates (Cramer
et al.,
2008; Kellar
et al.,
2013). The northeastern offshore and coastal stocks of pantropical spotted dolphins are likely to be encountered during the proposed surveys.
Spinner Dolphin
The spinner dolphin is pantropical in distribution, including oceanic tropical and sub-tropical waters between 40° N and 40° S (Jefferson
et al.,
2015). It is generally considered a pelagic species, but it can also be found in coastal waters (Perrin 2018b). In the ETP, three types of spinner dolphins have been identified and two of those are recognized as subspecies: The eastern spinner dolphin (
Stenella longirostris orientalis), considered an offshore species, the Central American spinner (
S. l. centroamericana;
also known as the Costa Rican spinner), considered a coastal species occurring from southern Mexico to Costa Rica (Perrin 1990; Dizon
et al.,
1991), and the ‘whitebelly’ spinner which is thought to be a hybrid of the eastern spinner and Gray's spinner (
S. l. longirostris). Gray's spinner dolphin is not expected to occur within the proposed study area. Although there is a great deal of overlap between the ranges of eastern and whitebelly spinner dolphins, the eastern form generally occurs in the northeastern portion of the ETP, whereas the whitebelly spinner occurs in the southern portion of the ETP, ranging farther offshore (Wade and Gerrodette 1993; Reilly and Fiedler 1994). Reilly and Fiedler (1994) noted that eastern spinners are associated with waters that have high surface temperatures and chlorophyll and shallow thermoclines, whereas whitebelly spinners are associated with cooler surface temperatures, lower chlorophyll levels, and deeper thermoclines. The eastern spinner dolphins are the most likely to occur in the proposed survey area (see Ferguson and Barlow 2001; Heckel
et al.,
2020), as this subspecies occurs in the ETP, east of 145° W, between 24° N off the Baja California Peninsula and 10° S off Peru (Perrin 1990). Wade and Gerrodette (1993) reported an abundance estimate of 1.7 million, and Gerrodette
et al.
(2005) estimated the abundance at 1.1 million for 2003. Gerrodette and Forcada (2005) noted that the population of eastern spinner dolphins had not yet recovered from the earlier population declines due to the tuna fishery. The population estimate for eastern spinner dolphins in 2003 was 612,662 (Gerrodette
et al.,
2005). In 2000, the whitebelly dolphin was estimated to number 801,000 in the ETP (Gerrodette
et al.,
2005). Bayliff (2004)
( printed page 19096)
noted a spinner dolphin mortality rate in the tuna fishery of 0.03 percent for 2004. Possible reasons why the population has not recovered include under-reported bycatch, effects of fishing activity on survival and reproduction, and long-term changes in the ecosystem (Gerrodette and Forcada, 2005). In 2008, Cramer
et al.,
determined that the continued chase and encirclement by the tuna fishery may be affecting the reproductive rates of the eastern spinner dolphin.
Guadalupe Fur Seal
During the summer season, most Guadalupe fur seal adults occur at rookeries in Mexico (Carretta et al. 2021). Most breeding and births occur at Isla Guadalupe, off the west coast of Baja California Peninsula; a secondary rookery exists at Isla Benito del Este (Maravilla-Chavez and Lowry 1999; Aurioles-Gamboa et al. 2010). Following the breeding season, adult males tend to move north to forage. All rookeries are outside of the project area as the proposed area is offshore. While at sea, this species is usually solitary but typically gathers in the hundreds to thousands at breeding sites. Guadalupe fur seals prefer rocky habitat for breeding and hauling out. They generally haul out at the base of towering cliffs on shores characterized by solid rock and large lava blocks (Peterson et al. 1968), although they can also inhabit caves and recesses (Belcher and Lee 2002). Guadalupe fur seals are unlikely to be encountered during the proposed seismic survey, as they typically occur farther north. However, Heckel et al. (2020) reported occasional records for Guerrero and Oaxaca.
Marine Mammal Hearing
Hearing is the most important sensory modality for marine mammals underwater, and exposure to anthropogenic sound can have deleterious effects. To appropriately assess the potential effects of exposure to sound, it is necessary to understand the frequency ranges marine mammals are able to hear. Not all marine mammal species have equal hearing capabilities (
e.g.,
Richardson
et al.,
1995; Wartzok and Ketten, 1999; Au and Hastings, 2008). To reflect this, Southall
et al.
(2007, 2019) recommended that marine mammals be divided into hearing groups based on directly measured (behavioral or auditory evoked potential techniques) or estimated hearing ranges (behavioral response data, anatomical modeling,
etc.). Generalized hearing ranges were chosen based on the ~65 dB threshold from composite audiograms, previous analyses in NMFS (2018), and/or data from Southall
et al.
(2007) and Southall
et al.
(2019). We note that the names of two hearing groups and the generalized hearing ranges of all marine mammal hearing groups have been recently updated (NMFS 2024) as reflected below in table 2.
Very High-frequency (VHF) cetaceans (true porpoises,
Kogia, river dolphins, Cephalorhynchid,
Lagenorhynchus cruciger & L. australis)
200 Hz to 165 kHz.
Phocid pinnipeds (PW) (underwater) (true seals)
40 Hz to 90 kHz.
Otariid pinnipeds (OW) (underwater) (sea lions and fur seals)
60 Hz to 68 kHz.
* Represents the generalized hearing range for the entire group as a composite (
i.e.,
all species within the group), where individual species' hearing ranges may not be as broad. Generalized hearing range chosen based on ~65 dB threshold from composite audiogram, previous analysis in NMFS 2018, and/or data from Southall et al. 2007; Southall
et al.
2019. Additionally, animals are able to detect very loud sounds above and below that “generalized” hearing range.
For more detail concerning these groups and associated frequency ranges, please see NMFS (2024) for a review of available information.
Potential Effects of Specified Activities on Marine Mammals and Their Habitat
This section provides a discussion of the ways in which components of the specified activity may impact marine mammals and their habitat. The Estimated Take of Marine Mammals section later in this document includes a quantitative analysis of the number of individuals that are expected to be taken by this activity. The Negligible Impact Analysis and Determination section considers the content of this section, the Estimated Take of Marine Mammals section, and the Proposed Mitigation section, to draw conclusions regarding the likely impacts of these activities on the reproductive success or survivorship of individuals and whether those impacts are reasonably expected to, or reasonably likely to, adversely affect the species or stock through effects on annual rates of recruitment or survival.
Description of Active Acoustic Sound Sources
This section contains a brief technical background on sound, the characteristics of certain sound types, and on metrics used in this proposal inasmuch as the information is relevant to the specified activity and to a discussion of the potential effects of the specified activity on marine mammals found later in this document.
Sound travels in waves, the basic components of which are frequency, wavelength, velocity, and amplitude. Frequency is the number of pressure waves that pass by a reference point per unit of time and is measured in hertz (Hz) or cycles per second. Wavelength is the distance between two peaks or corresponding points of a sound wave (length of one cycle). Higher frequency sounds have shorter wavelengths than lower frequency sounds, and typically attenuate (decrease) more rapidly, except in certain cases in shallower water. Amplitude is the height of the sound pressure wave or the “loudness” of a sound and is typically described using the relative unit of the dB. A sound pressure level (SPL) in dB is described as the ratio between a measured pressure and a reference pressure (for underwater sound, this is 1 micropascal (μPa)) and is a logarithmic unit that accounts for large variations in amplitude; therefore, a relatively small change in dB corresponds to large changes in sound pressure. The source level (SL) represents the SPL referenced at a distance of 1 m from the source (referenced to 1 μPa) while the received
( printed page 19097)
level is the SPL at the listener's position (referenced to 1 μPa).
Root mean square (RMS) is the quadratic mean sound pressure over the duration of an impulse. Root mean square is calculated by squaring all of the sound amplitudes, averaging the squares, and then taking the square root of the average (Urick, 1983). Root mean square accounts for both positive and negative values; squaring the pressures makes all values positive so that they may be accounted for in the summation of pressure levels (Hastings and Popper, 2005). This measurement is often used in the context of discussing behavioral effects, in part because behavioral effects, which often result from auditory cues, may be better expressed through averaged units than by peak pressures.
Sound exposure level (SEL; represented as dB re 1 μPa2
-s) represents the total energy contained within a pulse and considers both intensity and duration of exposure. Peak sound pressure (also referred to as zero-to-peak sound pressure or 0-p) is the maximum instantaneous sound pressure measurable in the water at a specified distance from the source and is represented in the same units as the RMS sound pressure. Another common metric is peak-to-peak sound pressure (pk-pk), which is the algebraic difference between the peak positive and peak negative sound pressures. Peak-to-peak pressure is typically approximately 6 dB higher than peak pressure (Southall
et al.,
2007).
When underwater objects vibrate or activity occurs, sound-pressure waves are created. These waves alternately compress and decompress the water as the sound wave travels. Underwater sound waves radiate in a manner similar to ripples on the surface of a pond and may be either directed in a beam or beams or may radiate in all directions (omnidirectional sources), as is the case for pulses produced by the airgun array considered here. The compressions and decompressions associated with sound waves are detected as changes in pressure by aquatic life and man-made sound receptors such as hydrophones.
Even in the absence of sound from the specified activity, the underwater environment is typically loud due to ambient sound. Ambient sound is defined as environmental background sound levels lacking a single source or point (Richardson
et al.,
1995), and the sound level of a region is defined by the total acoustical energy being generated by known and unknown sources. These sources may include physical (
e.g.,
wind and waves, earthquakes, ice, atmospheric sound), biological (
e.g.,
sounds produced by marine mammals, fish, and invertebrates), and anthropogenic (
e.g.,
vessels, dredging, construction) sound. A number of sources contribute to ambient sound, including the following (Richardson
et al.,
1995):
Wind and waves
—The complex interactions between wind and water surface, including processes such as breaking waves and wave-induced bubble oscillations and cavitation, are a main source of naturally occurring ambient sound for frequencies between 200 Hz and 50 kHz (Mitson, 1995). In general, ambient sound levels tend to increase with increasing wind speed and wave height. Surf sound becomes important near shore, with measurements collected at a distance of 8.5 km from shore showing an increase of 10 dB in the 100 to 700 Hz band during heavy surf conditions;
Precipitation
—Sound from rain and hail impacting the water surface can become an important component of total sound at frequencies above 500 Hz, and possibly down to 100 Hz during quiet times;
Biological
—Marine mammals can contribute significantly to ambient sound levels, as can some fish and snapping shrimp. The frequency band for biological contributions is from approximately 12 Hz to over 100 kHz; and
Anthropogenic
—Sources of anthropogenic sound related to human activity include transportation (surface vessels), dredging and construction, oil and gas drilling and production, seismic surveys, sonar, explosions, and ocean acoustic studies. Vessel noise typically dominates the total ambient sound for frequencies between 20 and 300 Hz. In general, the frequencies of anthropogenic sounds are below 1 kHz and, if higher frequency sound levels are created, they attenuate rapidly. Sound from identifiable anthropogenic sources other than the activity of interest (
e.g.,
a passing vessel) is sometimes termed background sound, as opposed to ambient sound.
The sum of the various natural and anthropogenic sound sources at any given location and time—which comprise “ambient” or “background” sound—depends not only on the source levels (as determined by current weather conditions and levels of biological and human activity) but also on the ability of sound to propagate through the environment. In turn, sound propagation is dependent on the spatially and temporally varying properties of the water column and sea floor, and is frequency-dependent. As a result of this dependence on a large number of varying factors, ambient sound levels can be expected to vary widely over both coarse and fine spatial and temporal scales. Sound levels at a given frequency and location can vary by 10-20 dB from day to day (Richardson
et al.,
1995). The result is that, depending on the source type and its intensity, sound from a given activity may be a negligible addition to the local environment or could form a distinctive signal that may affect marine mammals. Details of source types are described in the following text.
Sounds are often considered to fall into one of two general types: Pulsed and non-pulsed. The distinction between these two sound types is important because they have differing potential to cause physical effects, particularly with regard to hearing (
e.g.,
NMFS, 2018; Ward, 1997 in Southall
et al.,
2007). Please see Southall
et al.
(2007) for an in-depth discussion of these concepts.
Pulsed sound sources (
e.g.,
airguns, explosions, gunshots, sonic booms, impact pile driving) produce signals that are brief (typically considered to be less than one second), broadband, atonal transients (American National Standards Institute (ANSI), 1986, 2005; Harris, 1998; National Institute for Occupational Health and Safety (NIOSH), 1998; International Organization for Standardization (ISO), 2003) and occur either as isolated events or repeated in some succession. Pulsed sounds are all characterized by a relatively rapid rise from ambient pressure to a maximal pressure value followed by a rapid decay period that may include a period of diminishing, oscillating maximal and minimal pressures, and generally have an increased capacity to induce physical injury as compared with sounds that lack these features.
Non-pulsed sounds can be tonal, narrowband, or broadband, brief or prolonged, and may be either continuous or non-continuous (ANSI, 1995; NIOSH, 1998). Some of these non-pulsed sounds can be transient signals of short duration but without the essential properties of pulses (
e.g.,
rapid rise time). Examples of non-pulsed sounds include those produced by vessels, aircraft, machinery operations such as drilling or dredging, vibratory pile driving, and active sonar systems (such as those used by the U.S. Navy). The duration of such sounds, as received at a distance, can be greatly extended in a highly reverberant environment.
Airgun arrays produce pulsed signals with energy in a frequency range from about 10-2,000 Hz, with most energy radiated at frequencies below 200 Hz.
( printed page 19098)
The amplitude of the acoustic wave emitted from the source is equal in all directions (
i.e.,
omnidirectional), but airgun arrays do possess some directionality due to different phase delays between guns in different directions. Airgun arrays are typically tuned to maximize functionality for data acquisition purposes, meaning that sound transmitted in horizontal directions and at higher frequencies is minimized to the extent possible.
Acoustic Effects
Here, we discuss the effects of active acoustic sources on marine mammals.
Potential Effects of Underwater Sound[1]
—Anthropogenic sounds cover a broad range of frequencies and sound levels and can have a range of highly variable impacts on marine life, from none or minor to potentially severe responses, depending on received levels, duration of exposure, behavioral context, and various other factors. The potential effects of underwater sound from active acoustic sources can potentially result in one or more of the following: Temporary or permanent hearing impairment; non-auditory physical or physiological effects; behavioral disturbance; stress; and masking (Richardson
et al.,
1995; Gordon
et al.,
2004; Nowacek
et al.,
2007; Southall
et al.,
2007; Götz
et al.,
2009). The degree of effect is intrinsically related to the signal characteristics, received level, distance from the source, and duration of the sound exposure. In general, sudden, high level sounds can cause hearing loss, as can longer exposures to lower level sounds. Temporary or permanent loss of hearing, if it occurs at all, will occur almost exclusively in cases where a noise is within an animal's hearing frequency range. We first describe specific manifestations of acoustic effects before providing discussion specific to the use of airgun arrays.
Richardson
et al.
(1995) described zones of increasing intensity of effect that might be expected to occur, in relation to distance from a source and assuming that the signal is within an animal's hearing range. First is the area within which the acoustic signal would be audible (potentially perceived) to the animal, but not strong enough to elicit any overt behavioral or physiological response. The next zone corresponds with the area where the signal is audible to the animal and of sufficient intensity to elicit behavioral or physiological response. Third is a zone within which, for signals of high intensity, the received level is sufficient to potentially cause discomfort or tissue damage to auditory or other systems. Overlaying these zones to a certain extent is the area within which masking (
i.e.,
when a sound interferes with or masks the ability of an animal to detect a signal of interest that is above the absolute hearing threshold) may occur; the masking zone may be highly variable in size.
We describe the more severe effects of certain non-auditory physical or physiological effects only briefly as we do not expect that use of airgun arrays are reasonably likely to result in such effects (see below for further discussion). Potential effects from impulsive sound sources can range in severity from effects such as behavioral disturbance or tactile perception to physical discomfort, slight injury of the internal organs and the auditory system, or mortality (Yelverton
et al.,
1973). Non-auditory physiological effects or injuries that theoretically might occur in marine mammals exposed to high level underwater sound or as a secondary effect of extreme behavioral reactions (
e.g.,
change in dive profile as a result of an avoidance reaction) caused by exposure to sound include neurological effects, bubble formation, resonance effects, and other types of organ or tissue damage (Cox
et al.,
2006; Southall
et al.,
2007; Zimmer and Tyack, 2007; Tal
et al.,
2015). The survey activities considered here do not involve the use of devices such as explosives or mid-frequency tactical sonar that are associated with these types of effects.
Marine mammals, like all mammals, develop increased hearing thresholds over time due to age-related degeneration of auditory pathways and sensory cells of the inner ear. This natural, age-related hearing loss is contrasted by noise-induced hearing loss (Møller, 2012). Marine mammals exposed to high-intensity sound or to lower-intensity sound for prolonged periods can experience a noise-induced hearing threshold shift (TS), which NMFS defines as a change, usually an increase, in the threshold of audibility at a specified frequency or portion of an individual's hearing range above a previously established reference level as a result of noise exposure (NMFS, 2018, 2024). The amount of TS is customarily expressed in dB. Noise-induced hearing TS can be temporary (TTS) or permanent (PTS), and higher-level sound exposures are more likely to cause PTS or other AUD INJ. As described in NMFS (2018, 2024) there are numerous factors to consider when examining the consequence of TS, including, but not limited to, the signal temporal pattern (
e.g.,
impulsive or non-impulsive), likelihood an individual would be exposed for a long enough duration or to a high enough level to induce a TS, the magnitude of the TS, time to recovery (seconds to minutes or hours to days), the frequency range of the exposure (
i.e.,
spectral content), the hearing frequency range of the exposed species relative to the signal's frequency spectrum (
i.e.,
how animal uses sound within the frequency band of the signal;
e.g.,
Kastelein
et al.,
2014), and the overlap between the animal and the source (
e.g.,
spatial, temporal, and spectral).
Auditory Injury (AUD INJ)
NMFS (2024) defines AUD INJ as damage to the inner ear that can result in destruction of tissue, such as the loss of cochlear neuron synapses or auditory neuropathy (Houser 2021; Finneran 2024). AUD INJ may or may not result in a PTS. PTS is subsequently defined as a permanent, irreversible increase in the threshold of audibility at a specified frequency or portion of an individual's hearing range above a previously established reference level (NMFS, 2024). PTS does not generally affect more than a limited frequency range, and an animal that has incurred PTS has some level of hearing loss at the relevant frequencies; typically animals with PTS or other AUD INJ are not functionally deaf (Au and Hastings, 2008; Finneran, 2016). For marine mammals, AUD INJ is considered to be possible when sound exposures are sufficient to produce 40 dB of TTS measured after exposure (Southall
et al.
2007, 1019). AUD INJ levels for marine mammals are estimates, as with the exception of a single study unintentionally inducing PTS in a harbor seal (
Phoca vitulina) (Kastak
et al.,
2008; Reichmuth
et al.
2019), there are no empirical data measuring AUD INJ in marine mammals largely due to the fact that, for various ethical reasons, experiments involving anthropogenic noise exposure at levels inducing AUD INJ are not typically pursued or authorized (NMFS, 2024).
Temporary Threshold Shift (TTS)
TTS is the mildest form of hearing impairment that can occur during exposure to sound. TTS is a temporary, reversible increase in the threshold of audibility at a specified frequency or portion of an individual's hearing range above a previously established reference level (NMFS, 2024) that represents primarily tissue fatigue (Henderson
et al.,
2008), and is not considered an AUD INJ. Based on data from marine mammal
( printed page 19099)
TTS measurements (see Southall
et al.,
2007, 2019), a TTS of 6 dB is considered the minimum threshold shift clearly larger than any day-to-day or session-to-session variation in a subject's normal hearing ability (Finneran
et al.,
2000, 2002; Schlundt
et al.,
2000). While experiencing TTS, the hearing threshold rises, and a sound must be at a higher level in order to be heard.
In terrestrial and marine mammals, TTS can last from minutes or hours to days (
i.e.,
there is recovery back to baseline/pre-exposure levels), can occur within a specific frequency range (
i.e.,
an animal might only have a temporary loss of hearing sensitivity within a limited frequency band of its auditory range), and can be of varying amounts (
e.g.,
an animal's hearing sensitivity might be reduced by only 6 dB or reduced by 30 dB). In many cases, hearing sensitivity recovers rapidly after exposure to the sound ends. While there are data on sound levels and durations necessary to elicit mild TTS for marine mammals, recovery is complicated to predict and dependent on multiple factors.
Relationships between TTS and AUD INJ thresholds have not been studied in marine mammals, and there are no measured PTS data for cetaceans, but such relationships are assumed to be similar to those in humans and other terrestrial mammals. AUD INJ typically occurs at exposure levels at least several dB above that inducing mild TTS (
e.g.,
a 40-dB threshold shift approximates AUD INJ onset (Kryter
et al.,
1966; Miller, 1974), while a 6-dB threshold shift approximates TTS onset (Southall
et al.,
2007, 2019). Based on data from terrestrial mammals, a precautionary assumption is that the AUD INJ thresholds for impulsive sounds (such as airgun pulses as received close to the source) are at least 6 dB higher than the TTS threshold on a peak sound pressure level (PK SPL) basis and AUD INJ cumulative SEL (SEL24h) thresholds are 15 (impulsive sound criteria) to 20 dB (non-impulsive criteria) higher than TTS cumulative SEL thresholds (Southall
et al.,
2007, 2019). Given the higher level of sound or longer exposure duration necessary to cause AUD INJ as compared with TTS, it is considerably less likely that AUD INJ could occur.
Marine mammal hearing plays a critical role in communication with conspecifics, and interpretation of environmental cues for purposes such as predator avoidance and prey capture. Depending on the degree (elevation of threshold in dB), duration (
i.e.,
recovery time), and frequency range of TTS, and the context in which it is experienced, TTS can have effects on marine mammals ranging from discountable to serious. For example, a marine mammal may be able to readily compensate for a brief, relatively small amount of TTS in a non-critical frequency range that occurs during a time where ambient noise is lower and there are not as many competing sounds present. Alternatively, a larger amount and longer duration of TTS sustained during time when communication is critical for successful mother/calf interactions could have more serious impacts.
Finneran
et al.
(2015) measured hearing thresholds in 3 captive bottlenose dolphins before and after exposure to 10 pulses produced by a seismic airgun in order to study TTS induced after exposure to multiple pulses. Exposures began at relatively low levels and gradually increased over a period of several months, with the highest exposures at peak SPLs from 196 to 210 dB and cumulative (unweighted) SELs from 193-195 dB. No substantial TTS was observed. In addition, behavioral reactions were observed that indicated that animals can learn behaviors that effectively mitigate noise exposures (although exposure patterns must be learned, which is less likely in wild animals than for the captive animals considered in this study). The authors note that the failure to induce more significant auditory effects was likely due to the intermittent nature of exposure, the relatively low peak pressure produced by the acoustic source, and the low-frequency energy in airgun pulses as compared with the frequency range of best sensitivity for dolphins and other high-frequency cetaceans.
Currently, TTS data only exist for four species of cetaceans (bottlenose dolphin, beluga whale (
Delphinapterus leucas), harbor porpoise (
Phocoena phocoena), and Yangtze finless porpoise (
Neophocaena asiaeorientalis)) exposed to a limited number of sound sources (
i.e.,
mostly tones and octave-band noise) in laboratory settings (Finneran, 2015). In general, harbor porpoises have a lower TTS onset than other measured cetacean species (Finneran, 2015). Additionally, the existing marine mammal TTS data come from a limited number of individuals within these species.
Critical questions remain regarding the rate of TTS growth and recovery after exposure to intermittent noise and the effects of single and multiple pulses. Data at present are also insufficient to construct generalized models for recovery and determine the time necessary to treat subsequent exposures as independent events. More information is needed on the relationship between auditory evoked potential and behavioral measures of TTS for various stimuli. For summaries of data on TTS in marine mammals or for further discussion of TTS onset thresholds, please see Southall
et al.
(2007, 2019), Finneran and Jenkins (2012), Finneran (2015), and NMFS (2018, 2024).
Behavioral Effects
—Behavioral disturbance may include a variety of effects, including subtle changes in behavior (
e.g.,
minor or brief avoidance of an area or changes in vocalizations), more conspicuous changes in similar behavioral activities, and more sustained and/or potentially severe reactions, such as displacement from or abandonment of high-quality habitat. Behavioral responses to sound are highly variable and context-specific, and any reactions depend on numerous intrinsic and extrinsic factors (
e.g.,
species, state of maturity, experience, current activity, reproductive state, auditory sensitivity, time of day), as well as the interplay between factors (
e.g.,
Richardson
et al.,
1995; Wartzok
et al.,
2003; Southall
et al.,
2007, 2019; Weilgart, 2007; Archer
et al.,
2010). Behavioral reactions can vary not only among individuals but also within an individual, depending on previous experience with a sound source, context, and numerous other factors (Ellison
et al.,
2012), and can vary depending on characteristics associated with the sound source (
e.g.,
whether it is moving or stationary, number of sources, distance from the source). Please see appendices B-C of Southall
et al.
(2007) for a review of studies involving marine mammal behavioral responses to sound.
Habituation can occur when an animal's response to a stimulus wanes with repeated exposure, usually in the absence of unpleasant associated events (Wartzok
et al.,
2003). Animals are most likely to habituate to sounds that are predictable and unvarying. It is important to note that habituation is appropriately considered as a “progressive reduction in response to stimuli that are perceived as neither aversive nor beneficial,” rather than as, more generally, moderation in response to human disturbance (Bejder
et al.,
2009). The opposite process is sensitization, when an unpleasant experience leads to subsequent responses, often in the form of avoidance, at a lower level of exposure. As noted, behavioral state may affect the type of response. For example, animals that are resting may show greater behavioral change in response to disturbing sound levels than animals that are highly motivated to remain in
( printed page 19100)
an area for feeding (Richardson
et al.,
1995; National Research Council (NRC), 2003; Wartzok
et al.,
2003). Controlled experiments with captive marine mammals have shown pronounced behavioral reactions, including avoidance of loud sound sources (Ridgway
et al.,
1997). Observed responses of wild marine mammals to loud pulsed sound sources (typically seismic airguns or acoustic harassment devices) have been varied but often consist of avoidance behavior or other behavioral changes suggesting discomfort (Morton and Symonds, 2002; see also Richardson
et al.,
1995; Nowacek
et al.,
2007). However, many delphinids approach acoustic source vessels with no apparent discomfort or obvious behavioral change (
e.g.,
Barkaszi
et al.,
2012, Barkaszi and Kelly, 2018).
Available studies show wide variation in response to underwater sound; therefore, it is difficult to predict specifically how any given sound in a particular instance might affect marine mammals perceiving the signal. If a marine mammal does react briefly to an underwater sound by changing its behavior or moving a small distance, the impacts of the change are unlikely to be significant to the individual, let alone the stock or population. However, if a sound source displaces marine mammals from an important feeding or breeding area for a prolonged period, impacts on individuals and populations could be significant (
e.g.,
Lusseau and Bejder, 2007; Weilgart, 2007; NRC, 2005). There are broad categories of potential response, which we describe in greater detail here, that include alteration of dive behavior, alteration of foraging behavior, effects to breathing, interference with or alteration of vocalization, avoidance, and flight.
Changes in dive behavior can vary widely, and may consist of increased or decreased dive times and surface intervals as well as changes in the rates of ascent and descent during a dive (
e.g.,
Frankel and Clark, 2000; Ng and Leung, 2003; Nowacek
et al.,
2004; Goldbogen
et al.,
2013a, b). Variations in dive behavior may reflect disruptions in biologically significant activities (
e.g.,
foraging) or they may be of little biological significance. The impact of an alteration to dive behavior resulting from an acoustic exposure depends on what the animal is doing at the time of the exposure and the type and magnitude of the response.
Disruption of feeding behavior can be difficult to correlate with anthropogenic sound exposure, so it is usually inferred by observed displacement from known foraging areas, the appearance of secondary indicators (
e.g.,
bubble nets or sediment plumes), or changes in dive behavior. As for other types of behavioral response, the frequency, duration, and temporal pattern of signal presentation, as well as differences in species sensitivity, are likely contributing factors to differences in response in any given circumstance (
e.g.,
Croll
et al.,
2001; Nowacek
et al.;
2004; Madsen
et al.,
2006; Yazvenko
et al.,
2007a, b). A determination of whether foraging disruptions affect fitness consequences would require information on or estimates of the energetic requirements of the affected individuals and the relationship between prey availability, foraging effort and success, and the life history stage of the animal.
Visual tracking, passive acoustic monitoring (PAM), and movement recording tags were used to quantify sperm whale behavior prior to, during, and following exposure to airgun arrays at received levels in the range 140-160 dB at distances of 7-13 km, following a phase-in of sound intensity and full array exposures at 1-13 km (Madsen
et al.,
2006; Miller
et al.,
2009). Sperm whales did not exhibit horizontal avoidance behavior at the surface. However, foraging behavior may have been affected. The sperm whales exhibited 19 percent less vocal, or buzz, rate during full exposure relative to post exposure, and the whale that was approached most closely had an extended resting period and did not resume foraging until the airguns had ceased firing. The remaining whales continued to execute foraging dives throughout exposure; however, swimming movements during foraging dives were 6 percent lower during exposure than control periods (Miller
et al.,
2009). These data raise concerns that seismic surveys may impact foraging behavior in sperm whales, although more data are required to understand whether the differences were due to exposure or natural variation in sperm whale behavior (Miller
et al.,
2009).
Changes in respiration naturally vary with different behaviors and alterations to breathing rate as a function of acoustic exposure can be expected to co-occur with other behavioral reactions, such as a flight response or an alteration in diving. However, respiration rates in and of themselves may be representative of annoyance or an acute stress response. Various studies have shown that respiration rates may either be unaffected or could increase, depending on the species and signal characteristics, again highlighting the importance in understanding species differences in the tolerance of underwater noise when determining the potential for impacts resulting from anthropogenic sound exposure (
e.g.,
Kastelein
et al.,
2001, 2005, 2006; Gailey
et al.,
2007, 2016).
Marine mammals vocalize for different purposes and across multiple modes, such as whistling, echolocation click production, calling, and singing. Changes in vocalization behavior in response to anthropogenic noise can occur for any of these modes and may result from a need to compete with an increase in background noise or may reflect increased vigilance or a startle response. For example, in the presence of potentially masking signals, humpback whales and killer whales have been observed to increase the length of their songs or amplitude of calls (Miller
et al.,
2000; Fristrup
et al.,
2003; Foote
et al.,
2004; Holt
et al.,
2012), while right whales have been observed to shift the frequency content of their calls upward while reducing the rate of calling in areas of increased anthropogenic noise (Parks
et al.,
2007). In some cases, animals may cease sound production during production of aversive signals (Bowles
et al.,
1994).
Cerchio
et al.
(2014) used PAM to document the presence of singing humpback whales off the coast of northern Angola and to opportunistically test for the effect of seismic survey activity on the number of singing whales. Two recording units were deployed between March and December 2008 in the offshore environment; numbers of singers were counted every hour. Generalized additive mixed models were used to assess the effect of survey day (seasonality), hour (diel variation), moon phase, and received levels of noise (measured from a single pulse during each 10 minutes sampled period) on singer number. The number of singers significantly decreased with increasing received level of noise, suggesting that humpback whale communication was disrupted to some extent by the survey activity.
Castellote
et al.
(2012) reported acoustic and behavioral changes by fin whales in response to shipping and airgun noise. Acoustic features of fin whale song notes recorded in the Mediterranean Sea and northeast Atlantic Ocean were compared for areas with different shipping noise levels and traffic intensities and during a seismic airgun survey. During the first 72 hours of the survey, a steady decrease in song received levels and bearings to singers indicated that whales moved away from the acoustic source and out of the study area. This displacement persisted for a time period well beyond the 10-day duration of seismic airgun activity,
( printed page 19101)
providing evidence that fin whales may avoid an area for an extended period in the presence of increased noise. The authors hypothesize that fin whale acoustic communication is modified to compensate for increased background noise and that a sensitization process may play a role in the observed temporary displacement.
Seismic pulses at average received levels of 131 dB re 1 μPa2
-s caused blue whales to increase call production (Di Iorio and Clark, 2010). In contrast, McDonald
et al.
(1995) tracked a blue whale with seafloor seismometers and reported that it stopped vocalizing and changed its travel direction at a range of 10 km from the acoustic source vessel (estimated received level 143 dB pk-pk). Blackwell
et al.
(2013) found that bowhead whale call rates dropped significantly at onset of airgun use at sites with a median distance of 41-45 km from the survey. Blackwell
et al.
(2015) expanded this analysis to show that whales actually increased calling rates as soon as airgun signals were detectable before ultimately decreasing calling rates at higher received levels (
i.e.,
10-minute cumulative SEL (SELcum) of ~127 dB). Overall, these results suggest that bowhead whales may adjust their vocal output in an effort to compensate for noise before ceasing vocalization effort and ultimately deflecting from the acoustic source (Blackwell
et al.,
2013, 2015). These studies demonstrate that even low levels of noise received far from the source can induce changes in vocalization and/or behavior for mysticetes.
Avoidance is the displacement of an individual from an area or migration path as a result of the presence of sound or other stressors, and is one of the most obvious manifestations of disturbance in marine mammals (Richardson
et al.,
1995). For example, gray whales are known to change direction—deflecting from customary migratory paths—in order to avoid noise from seismic surveys (Malme
et al.,
1984). Humpback whales show avoidance behavior in the presence of an active seismic array during observational studies and controlled exposure experiments in western Australia (McCauley
et al.,
2000). Avoidance may be short-term, with animals returning to the area once the noise has ceased (
e.g.,
Bowles
et al.,
1994; Goold, 1996; Stone
et al.,
2000; Morton and Symonds, 2002; Gailey
et al.,
2007). Longer-term displacement is possible, however, which may lead to changes in abundance or distribution patterns of the affected species in the affected region if habituation to the presence of the sound does not occur (
e.g.,
Bejder
et al.,
2006; Teilmann
et al.,
2006).
Forney
et al.
(2017) detail the potential effects of noise on marine mammal populations with high site fidelity, including displacement and auditory masking, noting that a lack of observed response does not imply absence of fitness costs and that apparent tolerance of disturbance may have population-level impacts that are less obvious and difficult to document. Avoidance of overlap between disturbing noise and areas and/or times of particular importance for sensitive species may be critical to avoiding population-level impacts because (particularly for animals with high site fidelity) there may be a strong motivation to remain in the area despite negative impacts. Forney
et al.
(2017) state that, for these animals, remaining in a disturbed area may reflect a lack of alternatives rather than a lack of effects.
Forney
et al.
(2017) specifically discuss beaked whales, stating that until recently most knowledge of beaked whales was derived from strandings, as they have been involved in atypical mass stranding events associated with mid-frequency active (MFA) sonar training operations. Given these observations and recent research, beaked whales appear to be particularly sensitive and vulnerable to certain types of acoustic disturbance relative to most other marine mammal species. Individual beaked whales reacted strongly to experiments using simulated MFA sonar at low received levels, by moving away from the sound source and stopping foraging for extended periods. These responses, if on a frequent basis, could result in significant fitness costs to individuals (Forney
et al.,
2017). Additionally, difficulty in detection of beaked whales due to their cryptic surfacing behavior and silence when near the surface pose problems for mitigation measures employed to protect beaked whales. Forney
et al.
(2017) specifically states that failure to consider both displacement of beaked whales from their habitat and noise exposure could lead to more severe biological consequences.
A flight response is a dramatic change in normal movement to a directed and rapid movement away from the perceived location of a sound source. The flight response differs from other avoidance responses in the intensity of the response (
e.g.,
directed movement, rate of travel). Relatively little information on flight responses of marine mammals to anthropogenic signals exist, although observations of flight responses to the presence of predators have occurred (Connor and Heithaus, 1996). The result of a flight response could range from brief, temporary exertion and displacement from the area where the signal provokes flight to, in extreme cases, marine mammal strandings (Evans and England, 2001). However, it should be noted that response to a perceived predator does not necessarily invoke flight (Ford and Reeves, 2008), and whether individuals are solitary or in groups may influence the response.
Behavioral disturbance can also impact marine mammals in more subtle ways. Increased vigilance may result in costs related to diversion of focus and attention (
i.e.,
when a response consists of increased vigilance, it may come at the cost of decreased attention to other critical behaviors such as foraging or resting). These effects have generally not been demonstrated for marine mammals, but studies involving fish and terrestrial animals have shown that increased vigilance may substantially reduce feeding rates (
e.g.,
Beauchamp and Livoreil, 1997; Fritz
et al.,
2002; Purser and Radford, 2011). In addition, chronic disturbance can cause population declines through reduction of fitness (
e.g.,
decline in body condition) and subsequent reduction in reproductive success, survival, or both (
e.g.,
Harrington and Veitch, 1992; Daan
et al.,
1996; Bradshaw
et al.,
1998). However, Ridgway
et al.
(2006) reported that increased vigilance in bottlenose dolphins exposed to sound over a 5-day period did not cause any sleep deprivation or stress effects.
Many animals perform vital functions, such as feeding, resting, traveling, and socializing, on a diel cycle (24-hour cycle). Disruption of such functions resulting from reactions to stressors, such as sound exposure, are more likely to be significant if they last more than one diel cycle or recur on subsequent days (Southall
et al.,
2007). Consequently, a behavioral response lasting less than 1 day and not recurring on subsequent days is not considered particularly severe unless it could directly affect reproduction or survival (Southall
et al.,
2007). Note that there is a difference between multi-day substantive behavioral reactions and multi-day anthropogenic activities. For example, just because an activity lasts for multiple days does not necessarily mean that individual animals are either exposed to activity-related stressors for multiple days or, further, exposed in a manner resulting in sustained multi-day substantive behavioral responses.
Stone (2015) reported data from at-sea observations during 1,196 seismic surveys from 1994 to 2010. When large arrays of airguns (considered to be 500
( printed page 19102)
in3
(8,294 cc) or more in that study) were firing, lateral displacement, more localized avoidance, or other changes in behavior were evident for most odontocetes. However, significant responses to large arrays were found only for the minke whale and fin whale. Behavioral responses observed included changes in swimming or surfacing behavior, with indications that cetaceans remained near the water surface at these times. Cetaceans were recorded as feeding less often when large arrays were active. Behavioral observations of gray whales during a seismic survey monitored whale movements and respirations pre-, during, and post-seismic survey (Gailey
et al.,
2016). Behavioral state and water depth were the best “natural” predictors of whale movements and respiration and, after considering natural variation, none of the response variables were significantly associated with seismic survey or vessel sounds.
Stress Responses
—An animal's perception of a threat may be sufficient to trigger stress responses consisting of some combination of behavioral responses, autonomic nervous system responses, neuroendocrine responses, or immune responses (
e.g.,
Seyle, 1950; Moberg, 2000). In many cases, an animal's first and sometimes most economical (in terms of energetic costs) response is behavioral avoidance of the potential stressor. Autonomic nervous system responses to stress typically involve changes in heart rate, blood pressure, and gastrointestinal activity. These responses have a relatively short duration and may or may not have a significant long-term effect on an animal's fitness.
Neuroendocrine stress responses often involve the hypothalamus-pituitary-adrenal system. Virtually all neuroendocrine functions that are affected by stress—including immune competence, reproduction, metabolism, and behavior—are regulated by pituitary hormones. Stress-induced changes in the secretion of pituitary hormones have been implicated in failed reproduction, altered metabolism, reduced immune competence, and behavioral disturbance (
e.g.,
Moberg, 1987; Blecha, 2000). Increases in the circulation of glucocorticoids are also equated with stress (Romano
et al.,
2004).
The primary distinction between stress (which is adaptive and does not normally place an animal at risk) and distress is the cost of the response. During a stress response, an animal uses glycogen stores that can be quickly replenished once the stress is alleviated. In such circumstances, the cost of the stress response would not pose serious fitness consequences. However, when an animal does not have sufficient energy reserves to satisfy the energetic costs of a stress response, energy resources must be diverted from other functions. This state of distress will last until the animal replenishes its energetic reserves sufficiently to restore normal function.
Relationships between these physiological mechanisms, animal behavior, and the costs of stress responses are well-studied through controlled experiments and for both laboratory and free-ranging animals (
e.g.,
Holberton
et al.,
1996; Hood
et al.,
1998; Jessop
et al.,
2003; Krausman
et al.,
2004; Lankford
et al.,
2005). Stress responses due to exposure to anthropogenic sounds or other stressors and their effects on marine mammals have also been reviewed (Fair and Becker, 2000; Romano
et al.,
2002b) and, more rarely, studied in wild populations (
e.g.,
Romano
et al.,
2002a). For example, Rolland
et al.
(2012) found that noise reduction from reduced ship traffic in the Bay of Fundy was associated with decreased stress in North Atlantic right whales. These and other studies lead to a reasonable expectation that some marine mammals will experience physiological stress responses upon exposure to acoustic stressors and that it is possible that some of these would be classified as “distress.” In addition, any animal experiencing TTS would likely also experience stress responses (NRC, 2003).
Auditory Masking
—Sound can disrupt behavior through masking, or interfering with, an animal's ability to detect, recognize, or discriminate between acoustic signals of interest (
e.g.,
those used for intraspecific communication and social interactions, prey detection, predator avoidance, navigation) (Richardson
et al.,
1995; Erbe
et al.,
2016). Masking occurs when the receipt of a sound is interfered with by another coincident sound at similar frequencies and at similar or higher intensity, and may occur whether the sound is natural (
e.g.,
snapping shrimp, wind, waves, precipitation) or anthropogenic (
e.g.,
shipping, sonar, seismic exploration) in origin. The ability of a noise source to mask biologically important sounds depends on the characteristics of both the noise source and the signal of interest (
e.g.,
signal-to-noise ratio, temporal variability, direction), in relation to each other and to an animal's hearing abilities (
e.g.,
sensitivity, frequency range, critical ratios, frequency discrimination, directional discrimination, age or TTS hearing loss), and existing ambient noise and propagation conditions.
Under certain circumstances, significant masking could disrupt behavioral patterns, which in turn could affect fitness for survival and reproduction. It is important to distinguish TTS and PTS, which persist after the sound exposure, from masking, which occurs during the sound exposure. Because masking (without resulting in a TS) is not associated with abnormal physiological function, it is not considered a physiological effect, but rather a potential behavioral effect.
The frequency range of the potentially masking sound is important in predicting any potential behavioral impacts. For example, low-frequency signals may have less effect on high-frequency echolocation sounds produced by odontocetes but are more likely to affect detection of mysticete communication calls and other potentially important natural sounds such as those produced by surf and some prey species. The masking of communication signals by anthropogenic noise may be considered as a reduction in the communication space of animals (
e.g.,
Clark
et al.,
2009) and may result in energetic or other costs as animals change their vocalization behavior (
e.g.,
Miller
et al.,
2000; Foote
et al.,
2004; Parks
et al.,
2007; Di Iorio and Clark, 2010; Holt
et al.,
2009). Masking may be less in situations where the signal and noise come from different directions (Richardson
et al.,
1995), through amplitude modulation of the signal, or through other compensatory behaviors (Houser and Moore, 2014). Masking can be tested directly in captive species (
e.g.,
Erbe, 2008), but in wild populations it must be either modeled or inferred from evidence of masking compensation. There are few studies addressing real-world masking sounds likely to be experienced by marine mammals in the wild (
e.g.,
Branstetter
et al.,
2013).
Masking affects both senders and receivers of acoustic signals and can potentially have long-term chronic effects on marine mammals at the population level as well as at the individual level. Low-frequency ambient sound levels have increased by as much as 20 dB (more than three times in terms of SPL) in the world's ocean from pre-industrial periods, with most of the increase from distant commercial shipping (Hildebrand, 2009). All anthropogenic sound sources, but especially chronic and lower-frequency signals (
e.g.,
from vessel traffic), contribute to elevated ambient sound levels, thus intensifying masking.
( printed page 19103)
Masking effects of pulsed sounds (even from large arrays of airguns) on marine mammal calls and other natural sounds are expected to be limited, although there are few specific data on this. Because of the intermittent nature and low duty cycle of seismic pulses, animals can emit and receive sounds in the relatively quiet intervals between pulses. However, in exceptional situations, reverberation occurs for much or all of the interval between pulses (
e.g.,
Simard
et al.
2005; Clark and Gagnon 2006), which could mask calls. Situations with prolonged strong reverberation are infrequent. However, it is common for reverberation to cause some lesser degree of elevation of the background level between airgun pulses (
e.g.,
Gedamke 2011; Guerra
et al.
2011, 2016; Klinck
et al.
2012; Guan
et al.
2015), and this weaker reverberation presumably reduces the detection range of calls and other natural sounds to some degree. Guerra
et al.
(2016) reported that ambient noise levels between seismic pulses were elevated as a result of reverberation at ranges of 50 km from the seismic source. Based on measurements in deep water of the Southern Ocean, Gedamke (2011) estimated that the slight elevation of background noise levels during intervals between seismic pulses reduced blue and fin whale communication space by as much as 36-51 percent when a seismic survey was operating 450-2,800 km away. Based on preliminary modeling, Wittekind
et al.
(2016) reported that airgun sounds could reduce the communication range of blue and fin whales 2,000 km from the seismic source. Nieukirk
et al.
(2012) and Blackwell
et al.
(2013) noted the potential for masking effects from seismic surveys on large whales.
Some baleen and toothed whales are known to continue calling in the presence of seismic pulses, and their calls usually can be heard between the pulses (
e.g.,
Nieukirk
et al.
2012; Thode
et al.
2012; Bröker
et al.
2013; Sciacca
et al.
2016). Cerchio
et al.
(2014) suggested that the breeding display of humpback whales off Angola could be disrupted by seismic sounds, as singing activity declined with increasing received levels. In addition, some cetaceans are known to change their calling rates, shift their peak frequencies, or otherwise modify their vocal behavior in response to airgun sounds (
e.g.,
Di Iorio and Clark 2010; Castellote
et al.
2012; Blackwell
et al.
2013, 2015). The hearing systems of baleen whales are more sensitive to low-frequency sounds than are the ears of the small odontocetes that have been studied directly (
e.g.,
MacGillivray
et al.,
2014). The sounds important to small odontocetes are predominantly at much higher frequencies than are the dominant components of airgun sounds, thus limiting the potential for masking. In general, masking effects of seismic pulses are expected to be minor, given the normally intermittent nature of seismic pulses.
Vessel Noise
Vessel noise from the
Langseth
could affect marine animals in the proposed survey areas. Houghton
et al.
(2015) proposed that vessel speed is the most important predictor of received noise levels, and Putland
et al.
(2017) also reported reduced sound levels with decreased vessel speed. However, some energy is also produced at higher frequencies (Hermannsen
et al.,
2014); low levels of high-frequency sound from vessels has been shown to elicit responses in harbor porpoise (Dyndo
et al.,
2015).
Vessel noise, through masking, can reduce the effective communication distance of a marine mammal if the frequency of the sound source is close to that used by the animal, and if the sound is present for a significant fraction of time (
e.g.,
Richardson
et al.
1995; Clark
et al.,
2009; Jensen
et al.,
2009; Gervaise
et al.,
2012; Hatch
et al.,
2012; Rice
et al.,
2014; Dunlop 2015; Jones
et al.,
2017; Putland
et al.,
2017). In addition to the frequency and duration of the masking sound, the strength, temporal pattern, and location of the introduced sound also play a role in the extent of the masking (Branstetter
et al.,
2013, 2016; Finneran and Branstetter 2013; Sills
et al.,
2017). Branstetter
et al.
(2013) reported that time-domain metrics are also important in describing and predicting masking.
Baleen whales are thought to be more sensitive to sound at these low frequencies than are toothed whales (
e.g.,
MacGillivray
et al.
2014), possibly causing localized avoidance of the proposed survey area during seismic operations. Many odontocetes show considerable tolerance of vessel traffic, although they sometimes react at long distances if confined by ice or shallow water, if previously harassed by vessels, or have had little or no recent exposure to vessels (Richardson
et al.
1995). Pirotta
et al.
(2015) noted that the physical presence of vessels, not just ship noise, disturbed the foraging activity of bottlenose dolphins. There is little data on the behavioral reactions of beaked whales to vessel noise, though they seem to avoid approaching vessels (
e.g.,
Würsig
et al.,
1998) or dive for an extended period when approached by a vessel (
e.g.,
Kasuya, 1986).
In summary, project vessel sounds would not be at levels expected to cause anything more than possible localized and temporary behavioral changes in marine mammals, and would not be expected to result in significant negative effects on individuals or at the population level. In addition, in all oceans of the world, large vessel traffic is currently so prevalent that it is commonly considered a usual source of ambient sound (NSF-USGS, 2011).
Vessel Strike
Vessel collisions with marine mammals, or vessel strikes, can result in death or serious injury of the animal. Wounds resulting from vessel strike may include massive trauma, hemorrhaging, broken bones, or propeller lacerations (Knowlton and Kraus, 2001). An animal at the surface may be struck directly by a vessel, a surfacing animal may hit the bottom of a vessel, or an animal just below the surface may be cut by a vessel's propeller. Superficial strikes may not kill or result in the death of the animal. These interactions are typically associated with large whales (
e.g.,
fin whales), which are occasionally found draped across the bulbous bow of large commercial vessels upon arrival in port. Although smaller cetaceans are more maneuverable in relation to large vessels than are large whales, they may also be susceptible to strike. The severity of injuries typically depends on the size and speed of the vessel, with the probability of death or serious injury increasing as vessel speed increases (Knowlton and Kraus, 2001; Laist
et al.,
2001; Vanderlaan and Taggart, 2007; Conn and Silber, 2013). Impact forces increase with speed, as does the probability of a strike at a given distance (Silber
et al.,
2010; Gende
et al.,
2011).
Pace and Silber (2005) also found that the probability of death or serious injury increased rapidly with increasing vessel speed. Specifically, the predicted probability of serious injury or death increased from 45 to 75 percent as vessel speed increased from 10 to 14 knots (kn, 26 kilometer per hour (kph)), and exceeded 90 percent at 17 kn (31 kph). Higher speeds during collisions result in greater force of impact, but higher speeds also appear to increase the chance of severe injuries or death through increased likelihood of collision by pulling whales toward the vessel (Clyne, 1999; Knowlton
et al.,
1995). In a separate study, Vanderlaan and Taggart (2007) analyzed the probability of lethal mortality of large whales at a given speed, showing that
( printed page 19104)
the greatest rate of change in the probability of a lethal injury to a large whale as a function of vessel speed occurs between 8.6 and 15 kn (28 kph). The chances of a lethal injury decline from approximately 80 percent at 15 kn (28 kph) to approximately 20 percent at 8.6 kn (16 kph). At speeds below 11.8 kn (22 kph), the chances of lethal injury drop below 50 percent, while the probability asymptotically increases toward one hundred percent above 15 kn (28 kph).
The
Langseth
will travel at a speed of 5 kn (9 kph) while towing seismic survey gear. At this speed, both the possibility of striking a marine mammal and the possibility of a strike resulting in serious injury or mortality are discountable. At average transit speed, the probability of serious injury or mortality resulting from a strike is less than 50 percent. However, the likelihood of a strike actually happening is again discountable. Vessel strikes, as analyzed in the studies cited above, generally involve commercial shipping, which is much more common in both space and time than is geophysical survey activity. Jensen and Silber (2004) summarized vessel strikes of large whales worldwide from 1975-2003 and found that most collisions occurred in the open ocean and involved large vessels (
e.g.,
commercial shipping). No such incidents were reported for geophysical survey vessels during that time period.
It is possible for vessel strikes to occur while traveling at slow speeds. For example, a hydrographic survey vessel traveling at low speed (5.5 kn (10 kph)) while conducting mapping surveys off the central California coast struck and killed a blue whale in 2009. The State of California determined that the whale had suddenly and unexpectedly surfaced beneath the hull, with the result that the propeller severed the whale's vertebrae, and that this was an unavoidable event. This strike represents the only such incident in approximately 540,000 hours of similar coastal mapping activity (
p
= 1.9 × 10−6; 95 percent confidence interval = 0-5.5 × 10−6; NMFS, 2013). In addition, a research vessel reported a fatal strike in 2011 of a dolphin in the Atlantic, demonstrating that it is possible for strikes involving smaller cetaceans to occur. In that case, the incident report indicated that an animal apparently was struck by the vessel's propeller as it was intentionally swimming near the vessel. While indicative of the type of unusual events that cannot be ruled out, neither of these instances represents a circumstance that would be considered reasonably foreseeable or that would be considered preventable.
Although the likelihood of the vessel striking a marine mammal is low, we propose a robust vessel strike avoidance protocol (see Proposed Mitigation), which we believe eliminates any foreseeable risk of vessel strike during transit. We anticipate that vessel collisions involving a seismic data acquisition vessel towing gear, while not impossible, represent unlikely, unpredictable events for which there are no preventive measures. Given the proposed mitigation measures, the relatively slow speed of the vessel towing gear, the presence of bridge crew watching for obstacles at all times (including marine mammals), and the presence of marine mammal observers, the possibility of vessel strike is discountable and, further, were a strike of a large whale to occur, it would be unlikely to result in serious injury or mortality. No incidental take resulting from vessel strike is anticipated, and this potential effect of the specified activity will not be discussed further in the following analysis.
Stranding—
When a living or dead marine mammal swims or floats onto shore and becomes “beached” or incapable of returning to sea, the event is a “stranding” (Geraci
et al.,
1999; Perrin and Geraci, 2002; Geraci and Lounsbury, 2005; NMFS, 2007). The legal definition for a stranding under the MMPA is that a marine mammal is dead and is on a beach or shore of the United States; or in waters under the jurisdiction of the United States (including any navigable waters); or a marine mammal is alive and is on a beach or shore of the United States and is unable to return to the water; on a beach or shore of the United States and, although able to return to the water, is in need of apparent medical attention; or in the waters under the jurisdiction of the United States (including any navigable waters), but is unable to return to its natural habitat under its own power or without assistance.
Marine mammals strand for a variety of reasons, such as infectious agents, biotoxicosis, starvation, fishery interaction, vessel strike, unusual oceanographic or weather events, sound exposure, or combinations of these stressors sustained concurrently or in series. However, the cause or causes of most strandings are unknown (Geraci
et al.,
1976; Eaton, 1979; Odell
et al.,
1980; Best, 1982). Numerous studies suggest that the physiology, behavior, habitat relationships, age, or condition of cetaceans may cause them to strand or might predispose them to strand when exposed to another phenomenon. These suggestions are consistent with the conclusions of numerous other studies that have demonstrated that combinations of dissimilar stressors commonly combine to kill an animal or dramatically reduce its fitness, even though one exposure without the other does not produce the same result (Chroussos, 2000; Creel, 2005; DeVries
et al.,
2003; Fair and Becker, 2000; Foley
et al.,
2001; Moberg, 2000; Relyea, 2005a; 2005b, Romero, 2004; Sih
et al.,
2004).
There is no conclusive evidence that exposure to airgun noise results in behaviorally-mediated forms of injury. Behaviorally-mediated injury (
i.e.,
mass stranding events) has been primarily associated with beaked whales exposed to MFA sonar. MFA sonar and the alerting stimulus used in Nowacek
et al.
(2004) are very different from the noise produced by airguns. One should therefore not expect the same reaction to airgun noise as to these other sources. As explained below, military MFA sonar is very different from airguns, and one should not assume that airguns will cause the same effects as MFA sonar (including strandings).
To understand why military MFA sonar affects beaked whales differently than airguns do, it is important to note the distinction between behavioral sensitivity and susceptibility to auditory injury. To understand the potential for auditory injury in a particular marine mammal species in relation to a given acoustic signal, the frequency range the species is able to hear is critical, as well as the species' auditory sensitivity to frequencies within that range. Current data indicate that not all marine mammal species have equal hearing capabilities across all frequencies and, therefore, species are grouped into hearing groups with generalized hearing ranges assigned on the basis of available data (Southall
et al.,
2007, 2019). Hearing ranges as well as auditory sensitivity/susceptibility to frequencies within those ranges vary across the different groups. For example, in terms of hearing range, the very high-frequency cetaceans (
e.g., Kogia
spp.) have a generalized hearing range of frequencies between 200 Hz and 165 kHz, while high-frequency cetaceans—such as dolphins and beaked whales—have a generalized hearing range between 150 Hz to 160 kHz. Regarding auditory susceptibility within the hearing range, while high-frequency cetaceans and very high-frequency cetaceans have roughly similar hearing ranges, the high-frequency group is much more susceptible to noise-induced hearing loss during sound exposure,
i.e.,
these species have lower thresholds for
( printed page 19105)
these effects than other hearing groups (NMFS, 2018, 2024). Referring to a species as behaviorally sensitive to noise simply means that an animal of that species is more likely to respond to lower received levels of sound than an animal of another species that is considered less behaviorally sensitive. So, while dolphin species and beaked whale species—both in the high-frequency cetacean hearing group—are assumed to generally hear the same sounds equally well and be equally susceptible to noise-induced hearing loss (auditory injury), the best available information indicates that a beaked whale is more likely to behaviorally respond to that sound at a lower received level compared to an animal from other high-frequency cetacean species that are less behaviorally sensitive. This distinction is important because, while beaked whales are more likely to respond behaviorally to sounds than are many other species (even at lower levels), they cannot hear the predominant, lower frequency sounds from seismic airguns as well as sounds that have more energy at frequencies that beaked whales can hear better (such as military MFA sonar).
Military MFA sonar effects beaked whales differently than airguns do because it produces energy at different frequencies than airguns. High-frequency cetacean hearing is generically thought to be best between 8.8 to 110 kHz,
i.e.,
these cutoff values define the range above and below which a species in the group is assumed to have declining auditory sensitivity, until reaching frequencies that cannot be heard (NMFS, 2018, 2024). However, beaked whale hearing is likely best within a higher, narrower range (20-80 kHz, with best sensitivity around 40 kHz), based on a few measurements of hearing in stranded beaked whales (Cook
et al.,
2006; Finneran
et al.,
2009; Pacini
et al.,
2011) and several studies of acoustic signals produced by beaked whales (
e.g.,
Frantzis
et al.,
2002; Johnson
et al.,
2004, 2006; Zimmer
et al.,
2005). While precaution requires that the full range of audibility be considered when assessing risks associated with noise exposure (Southall
et al.,
2007, 2019), animals typically produce sound at frequencies where they hear best. More recently, Southall
et al.
(2019) suggested that certain species in the historical high-frequency hearing group (beaked whales, sperm whales, and killer whales) are likely more sensitive to lower frequencies within the group's generalized hearing range than are other species within the group, and state that the data for beaked whales suggest sensitivity to approximately 5 kHz. However, this information is consistent with the general conclusion that beaked whales (and other high-frequency cetaceans) are relatively insensitive to the frequencies where most energy of an airgun signal is found. Military MFA sonar is typically considered to operate in the frequency range of approximately 3-14 kHz (D'Amico
et al.,
2009),
i.e.,
outside the range of likely best hearing for beaked whales but within or close to the lower bounds, whereas most energy in an airgun signal is radiated at much lower frequencies, below 500 Hz (Dragoset, 1990).
It is important to distinguish between energy (loudness, measured in dB) and frequency (pitch, measured in Hz). In considering the potential impacts of mid-frequency components of airgun noise (1-10 kHz, where beaked whales can be expected to hear) on marine mammal hearing, one needs to account for the energy associated with these higher frequencies and determine what energy is truly “significant.” Although there is mid-frequency energy associated with airgun noise (as expected from a broadband source), airgun sound is predominantly below 1 kHz (Breitzke
et al.,
2008; Tashmukhambetov
et al.,
2008; Tolstoy
et al.,
2009). As stated by Richardson
et al.
(1995), “[. . .] most emitted [seismic airgun] energy is at 10-120 Hz, but the pulses contain some energy up to 500-1,000 Hz.” Tolstoy
et al.
(2009) conducted empirical measurements, demonstrating that sound energy levels associated with airguns were at least 20 dB lower at 1 kHz (considered “mid-frequency”) compared to higher energy levels associated with lower frequencies (below 300 Hz) (“all but a small fraction of the total energy being concentrated in the 10-300 Hz range” (Tolstoy
et al.,
2009), and at higher frequencies (
e.g.,
2.6-4 kHz), power might be less than 10 percent of the peak power at 10 Hz. Energy levels measured by Tolstoy
et al.
(2009) were even lower at frequencies above 1 kHz. In addition, as sound propagates away from the source, it tends to lose higher-frequency components faster than low-frequency components (
i.e.,
low-frequency sounds typically propagate longer distances than high-frequency sounds) (Diebold
et al.,
2010). Although higher-frequency components of airgun signals have been recorded, it is typically in surface-ducting conditions (
e.g.,
DeRuiter
et al.,
2006; Madsen
et al.,
2006) or in shallow water, where there are advantageous propagation conditions for the higher frequency (but low-energy) components of the airgun signal (Hermannsen
et al.,
2015). This should not be of concern because the likely behavioral reactions of beaked whales that can result in acute physical injury would result from noise exposure at depth (because of the potentially greater consequences of severe behavioral reactions). In summary, the frequency content of airgun signals is such that beaked whales will not be able to hear the signals well (compared to MFA sonar), especially at depth where we expect the consequences of noise exposure could be more severe.
Aside from frequency content, there are other significant differences between MFA sonar signals and the sounds produced by airguns that minimize the risk of severe behavioral reactions that could lead to strandings or deaths at sea,
e.g.,
significantly longer signal duration, horizontal sound direction, typical fast and unpredictable source movement. All of these characteristics of MFA sonar tend towards greater potential to cause severe behavioral or physiological reactions in exposed beaked whales that may contribute to stranding. Although both sources are powerful, MFA sonar contains significantly greater energy in the mid-frequency range, where beaked whales hear better. Short-duration, high energy pulses—such as those produced by airguns—have greater potential to cause damage to auditory structures (though this is unlikely for high-frequency cetaceans, as explained later in this document), but it is longer duration signals that have been implicated in the vast majority of beaked whale strandings. Faster, less predictable movements in combination with multiple source vessels are more likely to elicit a severe, potentially anti-predator response. Of additional interest in assessing the divergent characteristics of MFA sonar and airgun signals and their relative potential to cause stranding events or deaths at sea is the similarity between the MFA sonar signals and stereotyped calls of beaked whales' primary predator: the killer whale (Zimmer and Tyack, 2007). Although generic disturbance stimuli—as airgun noise may be considered in this case for beaked whales—may also trigger antipredator responses, stronger responses should generally be expected when perceived risk is greater, as when the stimulus is confused for a known predator (Frid and Dill, 2002). In addition, because the source of the perceived predator (
i.e.,
MFA sonar) will likely be closer to the whales (because attenuation limits the range of detection of mid-frequencies) and moving faster (because it will be on
( printed page 19106)
faster-moving vessels), any antipredator response would be more likely to be severe (with greater perceived predation risk, an animal is more likely to disregard the cost of the response; Frid and Dill, 2002). Indeed, when analyzing movements of a beaked whale exposed to playback of killer whale predation calls, Allen
et al.
(2014) found that the whale engaged in a prolonged, directed avoidance response, suggesting a behavioral reaction that could pose a risk factor for stranding. Overall, these significant differences between sound from MFA sonar and the mid-frequency sound component from airguns and the likelihood that MFA sonar signals will be interpreted in error as a predator are critical to understanding the likely risk of behaviorally-mediated injury due to seismic surveys.
The available scientific literature also provides a useful contrast between airgun noise and MFA sonar regarding the likely risk of behaviorally-mediated injury. There is strong evidence for the association of beaked whale stranding events with MFA sonar use, and particularly detailed accounting of several events is available (
e.g.,
a 2000 Bahamas stranding event for which investigators concluded that MFA sonar use was responsible; Evans and England, 2001). D'Amico
et al.,
(2009) reviewed 126 beaked whale mass stranding events over the period from 1950 (
i.e.,
from the development of modern MFA sonar systems) through 2004. Of these, there were two events where detailed information was available on both the timing and location of the stranding and the concurrent nearby naval activity, including verification of active MFA sonar usage, with no evidence for an alternative cause of stranding. An additional 10 events were at minimum spatially and temporally coincident with naval activity likely to have included MFA sonar use and, despite incomplete knowledge of timing and location of the stranding or the naval activity in some cases, there was no evidence for an alternative cause of stranding. The U.S. Navy has publicly stated agreement that five such events since 1996 were associated in time and space with MFA sonar use, either by the U.S. Navy alone or in joint training exercises with the North Atlantic Treaty Organization. The U.S. Navy additionally noted that, as of 2017, a 2014 beaked whale stranding event in Crete coincident with naval exercises was under review and had not yet been determined to be linked to sonar activities (U.S. Navy, 2017). Separately, the International Council for the Exploration of the Sea reported in 2005 that, worldwide, there have been about 50 known strandings, consisting mostly of beaked whales, with a potential causal link to MFA sonar (International Council for the Exploration for the Sea, 2005). In contrast, very few such associations have been made to seismic surveys, despite widespread use of airguns as a geophysical sound source in numerous locations around the world.
A review of possible stranding associations with seismic surveys (Castellote and Llorens, 2016) states that, “[s]peculation concerning possible links between seismic survey noise and cetacean strandings is available for a dozen events but without convincing causal evidence.” The authors' search of available information found 10 events worth further investigation via a ranking system representing a rough metric of the relative level of confidence offered by the data for inferences about the possible role of the seismic survey in a given stranding event. Only three of these events involved beaked whales. Whereas D'Amico
et al.,
(2009) used a 1-5 ranking system, in which “1” represented the most robust evidence connecting the event to MFA sonar use, Castellote and Llorens (2016) used a 1-6 ranking system, in which “6” represented the most robust evidence connecting the event to the seismic survey. As described above, D'Amico
et al.
(2009) found that two events were ranked “1” and 10 events were ranked “2” (
i.e.,
12 beaked whale stranding events were found to be associated with MFA sonar use). In contrast, Castellote and Llorens (2016) found that none of the three beaked whale stranding events achieved their highest ranks of 5 or 6. Of the 10 total events, none achieved the highest rank of 6. Two events were ranked as 5: one stranding in Peru involving dolphins and porpoises and a 2008 stranding in Madagascar. This latter ranking can only be broadly associated with the survey itself, as opposed to use of seismic airguns. An investigation of this stranding event, which did not involve beaked whales, concluded that use of a high-frequency mapping system (12-kHz multibeam echosounder) was the most plausible and likely initial behavioral trigger of the event, which was likely exacerbated by several site- and situation-specific secondary factors. The review panel found that seismic airguns were used after the initial strandings and animals entering a lagoon system, that airgun use clearly had no role as an initial trigger, and that there was no evidence that airgun use dissuaded animals from leaving (Southall
et al.,
2013).
However, one of these stranding events, involving two Cuvier's beaked whales, was contemporaneous with and reasonably associated spatially with a 2002 seismic survey in the Gulf of California conducted by L-DEO, as was the case for the 2007 Gulf of Cadiz seismic survey discussed by Castellote and Llorens (also involving two Cuvier's beaked whales). Neither event was considered a “true atypical mass stranding” (according to Frantzis (1998)) as used in the analysis of Castellote and Llorens (2016). While we agree with the authors that this lack of evidence should not be considered conclusive, it is clear that there is very little evidence that seismic surveys should be considered as posing a significant risk of acute harm to beaked whales or other high-frequency cetaceans. We have considered the potential for the proposed surveys to result in marine mammal stranding and, based on the best available information, do not expect a stranding to occur.
Entanglement
—Entanglements occur when marine mammals become wrapped around cables, lines, nets, or other objects suspended in the water column. During seismic operations, numerous cables, lines, and other objects primarily associated with the airgun array and hydrophone streamers will be towed behind the
Langseth
near the water's surface. However, we are not aware of any cases of entanglement of marine mammals in seismic survey equipment. No incidents of entanglement of marine mammals with seismic survey gear have been documented in over 54,000 nautical miles (100,000 km) of previous NSF-funded seismic surveys when observers were aboard (
e.g.,
Smultea and Holst 2003; Haley and Koski 2004; Holst 2004; Smultea
et al.,
2004; Holst
et al.,
2005; Haley and Ireland 2006; SIO and NSF 2006; Hauser
et al.,
2008; Holst and Smultea 2008). Although entanglement with the streamer is theoretically possible, it has not been documented during tens of thousands of miles of NSF-sponsored seismic cruises or, to our knowledge, during hundreds of thousands of miles of industrial seismic cruises. There are relatively few deployed devices, and no interaction between marine mammals and any such device has been recorded during prior NSF surveys using the devices. There are no meaningful entanglement risks posed by the proposed survey, and entanglement risks are not discussed further in this document.
( printed page 19107)
Anticipated Effects on Marine Mammal Habitat
Effects to Prey
—Marine mammal prey varies by species, season, and location and, for some, is not well documented. Fish react to sounds which are especially strong and/or intermittent low-frequency sounds, and behavioral responses such as flight or avoidance are the most likely effects. However, the reaction of fish to airguns depends on the physiological state of the fish, past exposures, motivation (
e.g.,
feeding, spawning, migration), and other environmental factors. Several studies have demonstrated that airgun sounds might affect the distribution and behavior of some fishes, potentially impacting foraging opportunities or increasing energetic costs (
e.g.,
Fewtrell and McCauley, 2012; Pearson
et al.,
1992; Skalski
et al.,
1992; Santulli
et al.,
1999; Paxton
et al.,
2017), though the bulk of studies indicate no or slight reaction to noise (
e.g.,
Miller and Cripps, 2013; Dalen and Knutsen, 1987; Peña
et al.,
2013; Chapman and Hawkins, 1969; Wardle
et al.,
2001; Jorgenson and Gyselman, 2009; Blaxter
et al.,
1981; Cott
et al.,
2012; Boeger
et al.,
2006), and that, most commonly, while there are likely to be impacts to fish as a result of noise from nearby airguns, such effects will be temporary. For example, investigators reported significant, short-term declines in commercial fishing catch rate of gadid fishes during and for up to 5 days after seismic survey operations, but the catch rate subsequently returned to normal (Engås
et al.,
1996; Engås and Lokkeborg, 2002). Other studies have reported similar findings (Hassel
et al.,
2004).
Skalski
et al.,
(1992) also found a reduction in catch rates—for rockfish (
Sebastes
spp.) in response to controlled airgun exposure—but suggested that the mechanism underlying the decline was not dispersal but rather decreased responsiveness to baited hooks associated with an alarm behavioral response. A companion study showed that alarm and startle responses were not sustained following the removal of the sound source (Pearson
et al.,
1992). Therefore, Skalski
et al.
(1992) suggested that the effects on fish abundance may be transitory, primarily occurring during the sound exposure itself. In some cases, effects on catch rates are variable within a study, which may be more broadly representative of temporary displacement of fish in response to airgun noise (
i.e.,
catch rates may increase in some locations and decrease in others) than any long-term damage to the fish themselves (Streever
et al.,
2016).
SPLs of sufficient strength have been known to cause injury to fish and fish mortality and, in some studies, fish auditory systems have been damaged by airgun noise (McCauley
et al.,
2003; Popper
et al.,
2005; Song
et al.,
2008). However, in most fish species, hair cells in the ear continuously regenerate and loss of auditory function likely is restored when damaged cells are replaced with new cells. Halvorsen
et al.
(2012) showed that a TTS of 4-6 dB was recoverable within 24 hours for one species. Impacts would be most severe when the individual fish is close to the source and when the duration of exposure is long; both of which are conditions unlikely to occur for this survey that is necessarily transient in any given location and likely result in brief, infrequent noise exposure to prey species in any given area. For this survey, the sound source is constantly moving, and most fish would likely avoid the sound source prior to receiving sound of sufficient intensity to cause physiological or anatomical damage. In addition, ramp-up may allow certain fish species the opportunity to move further away from the sound source.
A comprehensive review (Carroll
et al.,
2017) found that results are mixed as to the effects of airgun noise on the prey of marine mammals. While some studies suggest a change in prey distribution and/or a reduction in prey abundance following the use of seismic airguns, others suggest no effects or even positive effects in prey abundance. As one specific example, Paxton
et al.
(2017), which describes findings related to the effects of a 2014 seismic survey on a reef off of North Carolina, showed a 78 percent decrease in observed nighttime abundance for certain species. It is important to note that the evening hours during which the decline in fish habitat use was recorded (via video recording) occurred on the same day that the seismic survey passed, and no subsequent data is presented to support an inference that the response was long-lasting. Additionally, given that the finding is based on video images, the lack of recorded fish presence does not support a conclusion that the fish actually moved away from the site or suffered any serious impairment. In summary, this particular study corroborates prior studies indicating that a startle response or short-term displacement should be expected.
Available data suggest that cephalopods are capable of sensing the particle motion of sounds and detect low frequencies up to 1-1.5 kHz, depending on the species, and so are likely to detect airgun noise (Kaifu
et al.,
2008; Hu
et al.,
2009; Mooney
et al.,
2010; Samson
et al.,
2014). Auditory injuries (lesions occurring on the statocyst sensory hair cells) have been reported upon controlled exposure to low-frequency sounds, suggesting that cephalopods are particularly sensitive to low-frequency sound (André
et al.,
2011; Solé
et al.,
2013). Behavioral responses, such as inking and jetting, have also been reported upon exposure to low-frequency sound (McCauley
et al.,
2000b; Samson
et al.,
2014). Similar to fish, however, the transient nature of the survey leads to an expectation that effects will be largely limited to behavioral reactions and would occur as a result of brief, infrequent exposures.
With regard to potential impacts on zooplankton, McCauley
et al.
(2017) found that exposure to airgun noise resulted in significant depletion for more than half the taxa present and that there were two to three times more dead zooplankton after airgun exposure compared with controls for all taxa, within 1 km of the airguns. However, the authors also stated that in order to have significant impacts on r-selected species (
i.e.,
those with high growth rates and that produce many offspring) such as plankton, the spatial or temporal scale of impact must be large in comparison with the ecosystem concerned, and it is possible that the findings reflect avoidance by zooplankton rather than mortality (McCauley
et al.,
2017). In addition, the results of this study are inconsistent with a large body of research that generally finds limited spatial and temporal impacts to zooplankton as a result of exposure to airgun noise (
e.g.,
Dalen and Knutsen, 1987; Payne, 2004; Stanley
et al.,
2011). Most prior research on this topic, which has focused on relatively small spatial scales, has showed minimal effects (
e.g.,
Kostyuchenko, 1973; Booman
et al.,
1996; Sætre and Ona, 1996; Pearson
et al.,
1994; Bolle
et al.,
2012).
A modeling exercise was conducted as a follow-up to the McCauley
et al.
(2017) study (as recommended by McCauley
et al.), in order to assess the potential for impacts on ocean ecosystem dynamics and zooplankton population dynamics (Richardson
et al.,
2017). Richardson
et al.
(2017) found that for copepods with a short life cycle in a high-energy environment, a full-scale airgun survey would impact copepod abundance up to 3 days following the end of the survey, suggesting that effects such as those found by McCauley
et al.
(2017) would not be expected to be detectable
( printed page 19108)
downstream of the survey areas, either spatially or temporally.
Notably, a more recently described study produced results inconsistent with those of McCauley
et al.
(2017). Researchers conducted a field and laboratory study to assess if exposure to airgun noise affects mortality, predator escape response, or gene expression of the copepod
Calanus finmarchicus
(Fields
et al.,
2019). Immediate mortality of copepods was significantly higher, relative to controls, at distances of 5 m or less from the airguns. Mortality 1 week after the airgun blast was significantly higher in the copepods placed 10 m from the airgun but was not significantly different from the controls at a distance of 20 m from the airgun. The increase in mortality, relative to controls, did not exceed 30 percent at any distance from the airgun. Moreover, the authors caution that even this higher mortality in the immediate vicinity of the airguns may be more pronounced than what would be observed in free-swimming animals due to increased flow speed of fluid inside bags containing the experimental animals. There were no sublethal effects on the escape performance or the sensory threshold needed to initiate an escape response at any of the distances from the airgun that were tested. Whereas McCauley
et al.
(2017) reported an SEL of 156 dB at a range of 509-658 m, with zooplankton mortality observed at that range, Fields
et al.
(2019) reported an SEL of 186 dB at a range of 25 m, with no reported mortality at that distance. Regardless, if we assume a worst-case likelihood of severe impacts to zooplankton within approximately 1 km of the acoustic source, the brief time to regeneration of the potentially affected zooplankton populations does not lead us to expect any meaningful follow-on effects to the prey base for marine mammals.
A review article concluded that, while laboratory results provide scientific evidence for high-intensity and low-frequency sound-induced physical trauma and other negative effects on some fish and invertebrates, the sound exposure scenarios in some cases are not realistic to those encountered by marine organisms during routine seismic operations (Carroll
et al.,
2017). The review finds that there has been no evidence of reduced catch or abundance following seismic activities for invertebrates, and that there is conflicting evidence for fish with catch observed to increase, decrease, or remain the same. Further, where there is evidence for decreased catch rates in response to airgun noise, these findings provide no information about the underlying biological cause of catch rate reduction (Carroll
et al.,
2017).
In summary, impacts of the specified activity on marine mammal prey species will likely generally be limited to behavioral responses, the majority of prey species will be capable of moving out of the area during the survey, a rapid return to normal recruitment, distribution, and behavior for prey species is anticipated, and, overall, impacts to prey species will be minor and temporary. Prey species exposed to sound might move away from the sound source, experience TTS, experience masking of biologically relevant sounds, or show no obvious direct effects. Mortality from decompression injuries is possible in close proximity to a sound, but only limited data on mortality in response to airgun noise exposure are available (Hawkins
et al.,
2014). The most likely impacts for most prey species in the survey area would be temporary avoidance of the area. The proposed survey would move through an area relatively quickly, limiting exposure to multiple impulsive sounds. In all cases, sound levels would return to ambient once the survey moves out of the area or ends and the noise source is shut down and, when exposure to sound ends, behavioral and/or physiological responses are expected to end relatively quickly (McCauley
et al.,
2000). The duration of fish avoidance of a given area after survey effort stops is unknown, but a rapid return to normal recruitment, distribution, and behavior is anticipated. While the potential for disruption of spawning aggregations or schools of important prey species can be meaningful on a local scale, the mobile and temporary nature of this survey and the likelihood of temporary avoidance behavior suggest that impacts would be minor.
Acoustic Habitat
—Acoustic habitat is the soundscape—which encompasses all of the sound present in a particular location and time, as a whole—when considered from the perspective of the animals experiencing it. Animals produce sound for, or listen for sounds produced by, conspecifics (communication during feeding, mating, and other social activities), other animals (finding prey or avoiding predators), and the physical environment (finding suitable habitats, navigating). Together, sounds made by animals and the geophysical environment (
e.g.,
produced by earthquakes, lightning, wind, rain, waves) make up the natural contributions to the total acoustics of a place. These acoustic conditions, termed acoustic habitat, are one attribute of an animal's total habitat.
Soundscapes are also defined by, and acoustic habitat influenced by, the total contribution of anthropogenic sound. This may include incidental emissions from sources such as vessel traffic, or may be intentionally introduced to the marine environment for data acquisition purposes (as in the use of airgun arrays). Anthropogenic noise varies widely in its frequency content, duration, and loudness and these characteristics greatly influence the potential habitat-mediated effects to marine mammals (please see also the previous discussion on masking under
Acoustic Effects), which may range from local effects for brief periods of time to chronic effects over large areas and for long durations. Depending on the extent of effects to habitat, animals may alter their communications signals (thereby potentially expending additional energy) or miss acoustic cues (either conspecific or adventitious). For more detail on these concepts see,
e.g.,
Barber
et al.,
2010; Pijanowski
et al.,
2011; Francis and Barber, 2013; Lillis
et al.,
2014.
Problems arising from a failure to detect cues are more likely to occur when noise stimuli are chronic and overlap with biologically relevant cues used for communication, orientation, and predator/prey detection (Francis and Barber, 2013). Although the signals emitted by seismic airgun arrays are generally low frequency, they would also likely be of short duration and transient in any given area due to the nature of these surveys. As described previously, exploratory surveys such as these cover a large area but would be transient rather than focused in a given location over time and therefore would not be considered chronic in any given location.
Based on the information discussed herein, we conclude that impacts of the specified activity are not likely to have more than short-term adverse effects on any prey habitat or populations of prey species. Further, any impacts to marine mammal habitat are not expected to result in significant or long-term consequences for individual marine mammals, or to contribute to adverse impacts on their populations.
Estimated Take of Marine Mammals
This section provides an estimate of the number of incidental takes proposed for authorization through the IHA, which will inform NMFS' consideration of “small numbers,” the negligible impact determinations, and impacts on subsistence uses.
Harassment is the only type of take expected to result from these activities. Except with respect to certain activities
( printed page 19109)
not pertinent here, section 3(18) of the MMPA defines “harassment” as any act of pursuit, torment, or annoyance, which (i) has the potential to injure a marine mammal or marine mammal stock in the wild (Level A harassment); or (ii) has the potential to disturb a marine mammal or marine mammal stock in the wild by causing disruption of behavioral patterns, including, but not limited to, migration, breathing, nursing, breeding, feeding, or sheltering (Level B harassment).
Authorized takes would be by Level B harassment only, in the form of behavioral reactions and/or TTS, for individual marine mammals resulting from exposure to noise from the use of seismic airguns. Based on the nature of the activity and the anticipated effectiveness of the mitigation measures (
i.e.,
shutdown) discussed in detail below in the Proposed Mitigation section, Level A harassment is neither anticipated nor proposed to be authorized.
As described previously, no serious injury or mortality is anticipated or proposed to be authorized for this activity. Below we describe how the proposed take numbers are estimated.
For acoustic impacts, generally speaking, we estimate take by considering: (1) acoustic criteria above which NMFS believes the best available science indicates marine mammals will likely be behaviorally harassed or incur some degree of AUD INJ; (2) the area or volume of water that will be ensonified above these levels in a day; (3) the density or occurrence of marine mammals within these ensonified areas; and, (4) the number of days of activities. We note that while these factors can contribute to a basic calculation to provide an initial prediction of potential takes, additional information that can qualitatively inform take estimates is also sometimes available (
e.g.,
previous monitoring results or average group size). Below, we describe the factors considered here in more detail and present the proposed take estimates.
Acoustic Criteria
NMFS recommends the use of acoustic criteria that identify the received level of underwater sound above which exposed marine mammals would be reasonably expected to be behaviorally harassed (equated to Level B harassment) or to incur AUD INJ of some degree (equated to Level A harassment). We note that the criteria for AUD INJ, as well as the names of two hearing groups, have been recently updated (NMFS 2024) as reflected below in the Level A Harassment section.
Level B Harassment
—Though significantly driven by received level, the onset of behavioral disturbance from anthropogenic noise exposure is also informed to varying degrees by other factors related to the source or exposure context (
e.g.,
frequency, predictability, duty cycle, duration of the exposure, signal-to-noise ratio, distance to the source), the environment (
e.g.,
bathymetry, other noises in the area, predators in the area), and the receiving animals (hearing, motivation, experience, demography, life stage, depth) and can be difficult to predict (
e.g.,
Southall
et al.,
2007, 2021, Ellison
et al.,
2012). Based on what the available science indicates and the practical need to use a threshold based on a metric that is both predictable and measurable for most activities, NMFS typically uses a generalized acoustic threshold based on received level to estimate the onset of behavioral harassment. NMFS generally predicts that marine mammals are likely to be behaviorally harassed in a manner considered to be Level B harassment when exposed to underwater anthropogenic noise above root-mean-squared pressure received levels (RMS SPL) of 120 dB (referenced to 1 micropascal (re 1 μPa)) for continuous (
e.g.,
vibratory pile driving, drilling) and above RMS SPL 160 dB re 1 μPa for non-explosive impulsive (
e.g.,
seismic airguns) or intermittent (
e.g.,
scientific sonar) sources. Generally speaking, Level B harassment take estimates based on these behavioral harassment thresholds are expected to include any likely takes by TTS as, in most cases, the likelihood of TTS occurs at distances from the source less than those at which behavioral harassment is likely. TTS of a sufficient degree can manifest as behavioral harassment, as reduced hearing sensitivity and the potential reduced opportunities to detect important signals (conspecific communication, predators, prey) may result in changes in behavior patterns that would not otherwise occur.
L-DEO's proposed activity includes the use of impulsive seismic sources (
i.e.,
airguns), and therefore the RMS SPL thresholds of 160 dB re 1 μPa is applicable.
Level A harassment
—NMFS' Updated Technical Guidance for Assessing the Effects of Anthropogenic Sound on Marine Mammal Hearing (Version 3.0) (Updated Technical Guidance, 2024) identifies dual criteria to assess AUD INJ (Level A harassment) to five different underwater marine mammal groups (based on hearing sensitivity) as a result of exposure to noise from two different types of sources (impulsive or non-impulsive). L-DEO's proposed activity includes the use of impulsive seismic sources (
i.e.,
airguns).
* Dual metric criteria for impulsive sounds: Use whichever criteria results in the larger isopleth for calculating AUD INJ onset. If a non-impulsive sound has the potential of exceeding the peak SPL criteria associated with impulsive sounds, the PK SPL criteria are recommended for consideration for non-impulsive sources.
( printed page 19110)
Note:
Peak SPL (
Lp,0-pk
) has a reference value of 1 μPa, and weighted cumulative SEL (
LE,p
) has a reference value of 1 μPa2
s. In this table, criteria are abbreviated to be more reflective of International Organization for Standardization standards (ISO 2017; ISO 2020). The subscript “flat” is being included to indicate peak sound pressure are flat weighted or unweighted within the generalized hearing range of marine mammals underwater (
i.e.,
7 Hz to 165 kHz). The subscript associated with cumulative SEL criteria indicates the designated marine mammal auditory weighting function (LF, HF, and VHF cetaceans, and PW and OW pinnipeds) and that the recommended accumulation period is 24 hours. The
LE,p
criteria could be exceeded in a multitude of ways (
i.e.,
varying exposure levels and durations, duty cycle). When possible, it is valuable for action proponents to indicate the conditions under which these criteria will be exceeded.
Ensonified Area
Here, we describe operational and environmental parameters of the activity that are used in estimating the area ensonified above the acoustic thresholds, including source levels and transmission loss coefficient.
The ensonified area associated with Level A harassment is more technically challenging to predict due to the need to account for a duration component. Therefore, NMFS developed an optional User Spreadsheet tool to accompany the 2024 Updated Technical Guidance that can be used to relatively simply predict an isopleth distance for use in conjunction with marine mammal density or occurrence to help predict potential takes. We note that because of some of the assumptions included in the methods underlying this optional tool, we anticipate that the resulting isopleth estimates are typically going to be overestimates of some degree, which may result in an overestimate of potential take by Level A harassment. However, this optional tool offers the best way to estimate isopleth distances when more sophisticated modeling methods are not available or practical.
The proposed survey would entail the use of a cluster of two GI airguns with a total discharge volume of 90 in3
(1,475 cc) at a tow depth of 3 m. L-DEO model results are used to determine the 160 dB RMS radius for the airgun source down to a maximum depth of 2,000 m. Received sound levels have been predicted by L-DEO's model (Diebold
et al.
2010) as a function of distance from the airgun array. This modeling approach uses ray tracing for the direct wave traveling from the array to the receiver and its associated source ghost (reflection at the air-water interface in the vicinity of the array), in a constant-velocity half-space (infinite homogeneous ocean layer, unbounded by a seafloor). In addition, propagation measurements of pulses from the 36-airgun array at a tow depth of 6 m have been reported in deep water (~1,600 m), intermediate water depth on the slope (~600-1,100 m), and shallow water (~50 m) in the Gulf of America (previously Gulf of Mexico) (Tolstoy
et al.
2009; Diebold
et al.
2010).
For deep and intermediate water cases, the field measurements cannot be used readily to derive the harassment isopleths, as at those sites the calibration hydrophone was located at a roughly constant depth of 350-550 m, which may not intersect all the SPL isopleths at their widest point from the sea surface down to the assumed maximum relevant water depth (~2000 m) for marine mammals. At short ranges, where the direct arrivals dominate and the effects of seafloor interactions are minimal, the data at the deep sites are suitable for comparison with modeled levels at the depth of the calibration hydrophone. At longer ranges, the comparison with the model—constructed from the maximum SPL through the entire water column at varying distances from the airgun array—is the most relevant.
In deep and intermediate water depths at short ranges, sound levels for direct arrivals recorded by the calibration hydrophone and L-DEO model results for the same array tow depth are in good alignment (see figures 12 and 14 in Diebold
et al.
2010). Consequently, isopleths falling within this domain can be predicted reliably by the L-DEO model, although they may be imperfectly sampled by measurements recorded at a single depth. At greater distances, the calibration data show that seafloor-reflected and sub-seafloor-refracted arrivals dominate, whereas the direct arrivals become weak and/or incoherent (see figures 11, 12, and 16 in Diebold
et al.
2010). Aside from local topography effects, the region around the critical distance is where the observed levels rise closest to the model curve. However, the observed sound levels are found to fall almost entirely below the model curve. Thus, analysis of the Gulf of America calibration measurements demonstrates that although simple, the L-DEO model is a robust tool for conservatively estimating isopleths.
The proposed low-energy survey would acquire data with the 2 GI airgun array at a tow depth of 3 m. For deep water (>1,000 m), we use the deep-water radii obtained from the L-DEO model results down to a maximum water depth of 2,000 m for the airgun array.
L-DEO's modeling methodology is described in greater detail in their application. The estimated distances to the Level B harassment isopleth for the proposed airgun configuration are shown in table 4.
Table 4—Predicted Radial Distances From the
Langseth
Seismic Source to Isopleth Corresponding to Level B Harassment Threshold
Airgun configuration
Tow depth
(m)
Water depth
(m)
Predicted distances
(in m) to the Level B
harassment threshold
Two 45 in3
airguns
3
>1,000
438
Table 5—Modeled Radial Distance to Isopleths Corresponding to Level A Harassment Thresholds
Low
frequency
cetaceans
High
frequency
cetaceans
Very high
frequency
cetaceans
Phocid
pinnipeds
Otariid
pinnipeds
PTS SEL
cum
31.5
0
0.1
0.5
0
PTS Peak
3.9
1.1
35.3
3.4
1.1
The largest distance (in bold) of the dual criteria (SEL
cum
or Peak) was used to estimate threshold distances and potential takes by Level A harassment.
( printed page 19111)
Table 5 presents the modeled Level A harassment isopleths for each marine mammal hearing group based on L-DEO modeling incorporated in the companion user spreadsheet, for the low-energy surveys with the shortest shot interval (
i.e.,
greatest potential to cause PTS based on accumulated sound energy) (NMFS 2018, 2024).
Predicted distances to Level A harassment isopleths, which vary based on marine mammal hearing groups, were calculated based on modeling performed by L-DEO using the Nucleus software program and the NMFS user spreadsheet, described below. The acoustic thresholds for impulsive sounds contained in the NMFS Technical Guidance were presented as dual metric acoustic thresholds using both SELcum
and peak sound pressure metrics (NMFS, 2016). As dual metrics, NMFS considers onset of PTS (Level A harassment) to have occurred when either one of the two metrics is exceeded (
i.e.,
metric resulting in the largest isopleth). The SELcum
metric considers both level and duration of exposure, as well as auditory weighting functions by marine mammal hearing group.
The SELcum
for the 2 GI airgun array is derived from calculating the modified farfield signature. The farfield signature is often used as a theoretical representation of the source level. To compute the farfield signature, the source level is estimated at a large distance (right) below the array (
e.g.,
9 km), and this level is back projected mathematically to a notional distance of 1 m from the array's geometrical center. However, it has been recognized that the source level from the theoretical farfield signature is never physically achieved at the source when the source is an array of multiple airguns separated in space (Tolstoy
et al.,
2009). Near the source (at short ranges, distances <1 km), the pulses of sound pressure from each individual airgun in the source array do not stack constructively as they do for the theoretical farfield signature. The pulses from the different airguns spread out in time such that the source levels observed or modeled are the result of the summation of pulses from a few airguns, not the full array (Tolstoy
et al.,
2009). At larger distances, away from the source array center, sound pressure of all the airguns in the array stack coherently, but not within one time sample, resulting in smaller source levels (a few dB) than the source level derived from the farfield signature. Because the farfield signature does not take into account the large array effect near the source and is calculated as a point source, the farfield signature is not an appropriate measure of the sound source level for large arrays. See L-DEO's application for further detail on acoustic modeling.
In consideration of the received sound levels in the near-field as described above, we expect the potential for Level A harassment of any species to be de minimis, even before the likely moderating effects of aversion and/or other compensatory behaviors (
e.g.,
Nachtigall
et al.,
2018) are considered. We do not anticipate that auditory injury or Level A harassment is a likely outcome for any species and do not propose to authorize any take by Level A harassment for any species, given the very small modeled zones of injury for those species (estimated zones are less than 36 m for all species), in context of distributed source dynamics.
The Level B harassment estimates are based on a consideration of the number of marine mammals that could be within the area around the operating airgun array where received levels of sound ≥160 dB re 1 μPa RMS are predicted to occur. The estimated numbers are based on the densities (numbers per unit area) of marine mammals expected to occur in the area in the absence of seismic surveys. To the extent that marine mammals tend to move away from seismic sources before the sound level reaches the criterion level and tend not to approach an operating airgun array, these estimates likely overestimate the numbers actually exposed to the specified level of sound.
Marine Mammal Occurrence
In this section we provide information about the occurrence of marine mammals, including density or other relevant information which will inform the take calculations.
L-DEO used habitat-based stratified marine mammal densities for summer (July-December) for the ETP when available (Barlow
et al.,
2009), and densities for the ETP from NMFS (2015b) for all other species (See table 3 in L-DEO's application). Barlow
et al.
(2009) used data from 16 NMFS Southwest Fisheries Science Center (SWFSC) ship-based cetacean and ecosystem assessment surveys between 1986 and 2006 to develop habitat models to predict density for 15 cetacean species in the ETP. Model predictions were then used in standard line-transect formulae to estimate density for each transect segment for each survey year. Predicted densities for each year were smoothed with geospatial methods to obtain a continuous grid of density estimates for the surveyed area in the ETP. These annual grids were then averaged to obtain a composite grid that represents our best estimates of cetacean density over the past 20 years in the ETP. The models developed by Barlow
et al.
(2009) have been incorporated into a web-based geographic information system (GIS) software system developed by Duke University's Strategic Environmental Research and Development Program. The habitat-based density models consist of 100 km × 100 km grid cells. Densities in the grid cells that overlapped the survey area were averaged for each of the three water depth categories (shallow, intermediate, deep).
The NMFS SWFSC also developed density estimates for species in the ETP that may be affected by their own fisheries research activities (NMFS 2015a). These estimates were derived from abundance estimates using ship-based surveys of marine mammals in the ETP, as reported by Gerrodette
et al.
(2008). While the SWFSC developed volumetric density estimates (animals/km3
) to account for typical dive depth of each species (0-200 m and >200 m), L-DEO used the area density (animals/km2
) to represent expected density across all water depth strata.
For the sei whale, for which NMFS (2015) reported a density of zero, L-DEO used the spring density for Baja from U.S. Navy (2017). This was done because even though there is a modeled density of zero, we do expect there is some potential for sei whale to be in the project area during the proposed survey. No densities were available for Blainville's beaked whale, ginkgo-toothed beaked whale, Deraniyagala's beaked whale, or pygmy beaked whale so density for
Mesoplodon
species was used. For California sea lion only 10 percent of the density from the wider ETP was used to account for the fact that California sea lions typically do not occur in water greater than 1,000 m.
Take Estimation
Here we describe how the information provided above is synthesized to produce a quantitative estimate of the take that is reasonably likely to occur and proposed for authorization.
In order to estimate the number of marine mammals predicted to be exposed to sound levels that would result in Level A or Level B harassment, radial distances from the airgun array to the predicted isopleth corresponding to the Level A harassment and Level B harassment thresholds are calculated, as described above. Those radial distances were then used to calculate the area(s) around the airgun array predicted to be ensonified to sound levels that exceed the harassment thresholds. The distance
( printed page 19112)
for the 160-dB Level B harassment threshold and auditory injury (Level A harassment) thresholds (based on L-DEO model results) was used to draw a buffer around the area expected to be ensonified (
i.e.,
the survey area). The ensonified areas were then increased by 25 percent to account for potential delays, which is equivalent to adding 25 percent to the proposed line km to be surveyed. The density for each species was then multiplied by the daily ensonified areas (increased as described above) and then multiplied by the number of survey days (seven) to estimate potential takes (see appendix B of L-DEO's application for more information).
L-DEO assumed that their estimates of marine mammal exposures above harassment thresholds equate to take and requested authorization of those takes. Those estimates in turn form the basis for our proposed take authorization numbers. Based on the nature of the activity and the anticipated effectiveness of the mitigation measures (
i.e.,
shutdown) discussed in detail below in the Proposed Mitigation section, and due to no Level A harassment being calculated, Level A harassment is neither anticipated nor proposed to be authorized. Estimated exposures and proposed take numbers for authorization are shown in table 6.
Table 6—Estimated Take Proposed for Authorization
Species
Estimated takes
by Level B
harassment
Proposed takes
by Level B
harassment
Population
abundance
Percent of
population
Humpback whale 1
0
b
1
d
2,566
0.04
Minke whale
0
c
2
115
1.74
Bryde's whale
1
a
2
d
649
0.31
Fin whale
0
c
1
d
145
0.69
Sei whale
0
c
1
e
29,600
<0.01
Sperm whale
0
a
8
2,810
0.28
Cuvier's beaked whale
2
2
e
20,000
0.01
Longman's beaked whale
0
a
3
e
1,007
0.3
Mesoplodon beaked whales 2
1
a
12
e
25,300
0.01
Risso's dolphin
27
27
d
24,084
0.11
Rough-toothed dolphin
16
16
d
37,511
0.04
Bottlenose dolphin
62
62
d
61,536
0.1
Pantropical spotted dolphin
223
223
d
146,296
0.15
Spinner dolphin 3
222
222
d
186,906
0.12
Striped dolphin
57
a
6
d
128,867
0.05
Common dolphin
75
a
254
d
283,196
0.09
Fraser's dolphin
23
a
39
e
289,300
0.14
Short-finned pilot whales
7
a
18
d
3,348
0.54
Killer whale
1
a
5
d
852
0.59
False killer whale
3
a
11
e
39,600
0.03
Pygmy killer whale
3
a
28
e
38,900
0.07
Melon-headed whale
4
a
199
e
45,400
0.44
Kogia spp 4
0
a
2
e
11,200
0.02
Guadalupe fur seal
13
13
e
34,107
0.04
California sea lion
3
3
e
105,000
<0.01
1
Takes are assumed to be from the Central America/Southern Mexico Stock.
2
Includes: Blainville's, ginkgo-toothed, Deraniyagala's, and pygmy beaked whales.
3
Includes both whitebelly and eastern population.
4
Includes pygmy and dwarf sperm whales.
a
Increased to a group size from Wade and Gerrodette (1993).
b
Increased to a group size from Oedekoven
et al.,
2021.
c
Increased to a group size based on 87 FR 27111 (May 6, 2022).
d
Population sizes are for Pacific waters of Mexico (Gerrodette and Palacios, 1996).
e
Population in ETP or wider Pacific (NMFS 2015).
Proposed Mitigation
In order to issue an IHA under section 101(a)(5)(D) of the MMPA, NMFS must set forth the permissible methods of taking pursuant to the activity, and other means of effecting the least practicable impact on the species or stock and its habitat, paying particular attention to rookeries, mating grounds, and areas of similar significance, and on the availability of the species or stock for taking for certain subsistence uses (latter not applicable for this action). NMFS regulations require applicants for incidental take authorizations to include information about the availability and feasibility (economic and technological) of equipment, methods, and manner of conducting the activity or other means of effecting the least practicable adverse impact upon the affected species or stocks, and their habitat (50 CFR 216.104(a)(11)).
In evaluating how mitigation may or may not be appropriate to ensure the least practicable adverse impact on species or stocks and their habitat, as well as subsistence uses where applicable, NMFS considers two primary factors:
(1) The manner in which, and the degree to which, the successful implementation of the measure(s) is expected to reduce impacts to marine mammals, marine mammal species or stocks, and their habitat. This considers the nature of the potential adverse impact being mitigated (likelihood, scope, range). It further considers the likelihood that the measure will be effective if implemented (probability of accomplishing the mitigating result if implemented as planned), the likelihood of effective implementation (probability implemented as planned), and;
(2) The practicability of the measures for applicant implementation, which may consider such things as cost, and impact on operations.
( printed page 19113)
Vessel-Based Visual Mitigation Monitoring
Visual monitoring requires the use of trained observers (herein referred to as visual protected species observers (PSOs)) to scan the ocean surface for the presence of marine mammals. The area to be scanned visually includes primarily the shutdown zone (SZ), within which observation of certain marine mammals requires shutdown of the acoustic source, a buffer zone, and to the extent possible depending on conditions, the surrounding waters. The buffer zone means an area beyond the SZ to be monitored for the presence of marine mammals that may enter the SZ. During pre-start clearance monitoring (
i.e.,
before ramp-up begins), the buffer zone also acts as an extension of the SZ in that observations of marine mammals within the buffer zone would also prevent airgun operations from beginning (
i.e.,
ramp-up). The buffer zone encompasses the area at and below the sea surface from the edge of the 0-100 m SZ, out to a radius of 200 m from the edges of the airgun array (100-200 m). This 200-m zone (SZ plus buffer) represents the pre-start clearance zone. Visual monitoring of the SZ and adjacent waters (buffer plus surrounding waters) is intended to establish and, when visual conditions allow, maintain zones around the sound source that are clear of marine mammals, thereby reducing or eliminating the potential for injury and minimizing the potential for more severe behavioral reactions for animals occurring closer to the vessel. Visual monitoring of the buffer zone is intended to (1) provide additional protection to marine mammals that may be in the vicinity of the vessel during pre-start clearance, and (2) during airgun use, aid in establishing and maintaining the SZ by alerting the visual observer and crew of marine mammals that are outside of, but may approach and enter, the SZ.
During survey operations (
e.g.,
any day on which use of the airgun array is planned to occur and whenever the airgun array is in the water, whether activated or not), a minimum of two visual PSOs must be on duty and conducting visual observations at all times during daylight hours (
i.e.,
from 30 minutes prior to sunrise through 30 minutes following sunset). Visual monitoring of the pre-start clearance zone must begin no less than 30 minutes prior to ramp-up and monitoring must continue until 1 hour after use of the airgun array ceases or until 30 minutes past sunset. Visual PSOs shall coordinate to ensure 360° visual coverage around the vessel from the most appropriate observation posts and shall conduct visual observations using binoculars and the naked eye while free from distractions and in a consistent, systematic, and diligent manner.
PSOs shall establish and monitor the SZ and buffer zone. These zones shall be based upon the radial distance from the edges of the airgun array (rather than being based on the center of the array or around the vessel itself). During use of the airgun array (
i.e.,
anytime airguns are active, including ramp-up), detections of marine mammals within the buffer zone (but outside the SZ) shall be communicated to the operator to prepare for the potential shutdown of the airgun array. Any observations of marine mammals by crew members shall be relayed to the PSO team. During good conditions (
e.g.,
daylight hours; Beaufort sea state (BSS) 3 or less), visual PSOs shall conduct observations when the airgun array is not operating for comparison of sighting rates and behavior with and without use of the airgun array and between acquisition periods, to the maximum extent practicable.
Visual PSOs may be on watch for a maximum of 4 consecutive hours followed by a break of at least 1 hour between watches and may conduct a maximum of 12 hours of observation per 24-hour period.
Establishment of Shutdown and Pre-Start Clearance Zones
A SZ is a defined area within which occurrence of a marine mammal triggers mitigation action intended to reduce the potential for certain outcomes (
e.g.,
auditory injury, disruption of critical behaviors). The PSOs would establish a minimum SZ with a 100-m radius. The 100-m SZ would be based on radial distance from the edge of the airgun array (rather than being based on the center of the array or around the vessel itself). With certain exceptions (described below), if a marine mammal appears within or enters this zone, the airgun array would be shut down.
The pre-start clearance zone is defined as the area that must be clear of marine mammals prior to beginning ramp-up of the airgun array and includes the SZ plus the buffer zone. Detections of marine mammals within the pre-start clearance zone would prevent airgun operations from beginning (
i.e.,
ramp-up).
The 100-m SZ is intended to be precautionary in the sense that it would be expected to contain sound exceeding the injury criteria for all cetacean hearing groups, (based on the dual criteria of SELcum
and peak SPL), while also providing a consistent, reasonably observable zone within which PSOs would typically be able to conduct effective observational effort. Additionally, a 100-m SZ is expected to minimize the likelihood that marine mammals will be exposed to levels likely to result in more severe behavioral responses. Although significantly greater distances may be observed from an elevated platform under good conditions, we expect that 100 m is likely regularly attainable for PSOs using the naked eye during typical conditions. The pre-start clearance zone simply represents the addition of a buffer to the SZ, doubling the SZ size during pre-clearance.
An extended SZ of 500 m must be implemented for all beaked whales, a large whale with a calf, and groups of six or more large whales. No buffer of this extended SZ is required, as NMFS concludes that this extended SZ is sufficiently protective to mitigate harassment to these groups.
Pre-Start Clearance and Ramp-Up
Ramp-up (sometimes referred to as “soft start”) means the gradual and systematic increase of emitted sound levels from an airgun array. The intent of pre-start clearance observation (30 minutes) is to ensure no marine mammals are observed within the pre-start clearance zone (or extended SZ, for beaked whales, a large whale with a calf, and groups of six or more large whales) prior to the beginning of ramp-up. During the pre-start clearance period is the only time observations of marine mammals in the buffer zone would prevent operations (
i.e.,
the beginning of ramp-up). The intent of the ramp-up is to warn marine mammals of pending seismic survey operations and to allow sufficient time for those animals to leave the immediate vicinity prior to the sound source reaching full intensity. A ramp-up procedure, involving a stepwise increase in the number of airguns firing and total array volume until all operational airguns are activated and the full volume is achieved, is required at all times as part of the activation of the airgun array. All operators must adhere to the following pre-start clearance and ramp-up requirements:
The operator must notify a designated PSO of the planned start of ramp-up as agreed upon with the lead PSO; the notification time should not be less than 60 minutes prior to the planned ramp-up in order to allow the PSOs time to monitor the pre-start clearance zone (and extended SZ) for 30 minutes prior to the initiation of ramp-up (pre-start clearance);
( printed page 19114)
Ramp-ups shall be scheduled so as to minimize the time spent with the source activated prior to reaching the designated run-in;
One of the PSOs conducting pre-start clearance observations must be notified again immediately prior to initiating ramp-up procedures and the operator must receive confirmation from the PSO to proceed;
Ramp-up may not be initiated if any marine mammal is within the applicable shutdown or buffer zone. If a marine mammal is observed within the pre-start clearance zone (or extended SZ, for beaked whales, a large whale with a calf, and groups of six or more large whales) during the 30 minute pre-start clearance period, ramp-up may not begin until the animal(s) has been observed exiting the zones or until an additional time period has elapsed with no further sightings (15 minutes for small odontocetes, and 30 minutes for all mysticetes and all other odontocetes, including sperm whales, beaked whales, and large delphinids, such as pilot whales);
Ramp-up must begin by activating one GI airgun followed by the second, with each stage lasting no less than 5 minutes. The operator must provide information to the PSO documenting that appropriate procedures were followed;
PSOs must monitor the pre-start clearance zone and extended SZ during ramp-up, and ramp-up must cease and the source must be shut down upon detection of a marine mammal within the applicable zone. Once ramp-up has begun, detections of marine mammals within the buffer zone do not require shutdown, but such observation shall be communicated to the operator to prepare for the potential shutdown;
Ramp-up may occur at times of poor visibility, including nighttime, if appropriate monitoring has occurred with no observations in the 30 minutes prior to beginning ramp-up. Airgun array activation may only occur at times of poor visibility where operational planning cannot reasonably avoid such circumstances;
If the airgun array is shut down for brief periods (i.e.,
less than 30 minutes) for reasons other than implementation of prescribed mitigation (
e.g.,
mechanical difficulty), it may be activated again without ramp-up if PSOs have maintained constant visual observation and no visual observations of marine mammals have occurred within the pre-start clearance zone (or extended SZ, where applicable). For any longer shutdown, pre-start clearance observation and ramp-up are required; and
Testing of the airgun array involving all elements requires ramp-up. Testing limited to individual source elements or strings does not require ramp-up but does require pre-start clearance of 30 minutes.
Shutdown
The shutdown of an airgun array requires the immediate de-activation of all individual airgun elements of the array. Any PSO on duty will have the authority to call for shutdown of the airgun array if a marine mammal is detected within the applicable SZ. The operator must also establish and maintain clear lines of communication directly between PSOs on duty and crew controlling the airgun array to ensure that shutdown commands are conveyed swiftly while allowing PSOs to maintain watch. When the airgun array is active (
i.e.,
anytime one or more airguns is active, including during ramp-up) and a marine mammal appears within or enters the applicable SZ the airgun array will be shut down. When shutdown is called for by a PSO, the airgun array will be immediately deactivated and any dispute resolved only following deactivation.
Following a shutdown, airgun activity would not resume until the marine mammal has cleared the SZ. The animal would be considered to have cleared the SZ if it is visually observed to have departed the SZ (
i.e.,
animal is not required to fully exit the buffer zone where applicable), or it has not been seen within the SZ for 15 minutes for small odontocetes or 30 minutes for all mysticetes and all other odontocetes, including sperm whales, beaked whales, and large delphinids, such as pilot whales.
The shutdown requirement is waived for specific genera of small dolphins if an individual is detected within the SZ. The small dolphin group is intended to encompass those members of the Family Delphinidae most likely to voluntarily approach the source vessel for purposes of interacting with the vessel and/or airgun array (
e.g.,
bow riding). This exception to the shutdown requirement applies solely to the specific genera of small dolphins (
Lagenodelphis, Stenella, Steno, Delphinus, Tursiops,
and pinnipeds).
We include this small dolphin exception because shutdown requirements for these species under all circumstances represent practicability concerns without likely commensurate benefits for the animals in question. Small dolphins are generally the most commonly observed marine mammals in the specific geographic region and would typically be the only marine mammals likely to intentionally approach the vessel. As described above, auditory injury is extremely unlikely to occur for high-frequency cetaceans (
e.g.,
delphinids), as this group is relatively insensitive to sound produced at the predominant frequencies in an airgun pulse while also having a relatively high threshold for the onset of auditory injury (
i.e.,
permanent threshold shift).
A large body of anecdotal evidence indicates that small dolphins commonly approach vessels and/or towed arrays during active sound production for purposes of bow riding with no apparent effect observed (
e.g.,
Barkaszi
et al.,
2012; Barkaszi and Kelly, 2018). The potential for increased shutdowns resulting from such a measure would require the
Langseth
to revisit the missed track line to reacquire data, resulting in an overall increase in the total sound energy input to the marine environment and an increase in the total duration over which the survey is active in a given area. Although other high-frequency hearing specialists (
e.g.,
large delphinids) are no more likely to incur auditory injury than are small dolphins, they are much less likely to approach vessels. Therefore, retaining a shutdown requirement for large delphinids would not have similar impacts in terms of either practicability for the applicant or corollary increase in sound energy output and time on the water. We do anticipate some benefit for a shutdown requirement for large delphinids in that it simplifies somewhat the total range of decision-making for PSOs and may preclude any potential for physiological effects other than to the auditory system as well as some more severe behavioral reactions for any such animals in close proximity to the
Langseth.
Visual PSOs shall use best professional judgment in making the decision to call for a shutdown if there is uncertainty regarding identification (
i.e.,
whether the observed marine mammal(s) belongs to one of the delphinid genera for which shutdown is waived or one of the species with a larger SZ).
L-DEO must implement shutdown if a marine mammal species for which take was not authorized or a species for which authorization was granted but the authorized takes have been met approaches the Level A or Level B harassment zones. L-DEO must also implement shutdown if any large whale (defined as a sperm whale or any mysticete species) with a calf (defined as an animal less than two-thirds the body size of an adult observed to be in close association with an adult) and/or
( printed page 19115)
an aggregation of six or more large whales are observed at any distance.
Vessel Strike Avoidance Mitigation Measures
Vessel personnel should use an appropriate reference guide that includes identifying information on all marine mammals that may be encountered. Vessel operators must comply with the below measures except under extraordinary circumstances when the safety of the vessel or crew is in doubt or the safety of life at sea is in question. These requirements do not apply in any case where compliance would create an imminent and serious threat to a person or vessel or to the extent that a vessel is restricted in its ability to maneuver and, because of the restriction, cannot comply.
Vessel operators and crews must maintain a vigilant watch for all marine mammals and slow down, stop their vessel, or alter course, as appropriate and regardless of vessel size, to avoid striking any marine mammal. A single marine mammal at the surface may indicate the presence of submerged animals in the vicinity of the vessel; therefore, precautionary measures should always be exercised. A visual observer aboard the vessel must monitor a vessel strike avoidance zone around the vessel (separation distances stated below). Visual observers monitoring the vessel strike avoidance zone may be third-party observers (
i.e.,
PSOs) or crew members, but crew members responsible for these duties must be provided sufficient training to (1) distinguish marine mammals from other phenomena and (2) broadly to identify a marine mammal as a large whale (defined in this context as sperm whales or baleen whales), or other marine mammals.
Vessel speeds must be reduced to 10 kn (18.5 kph) or less when mother/calf pairs, pods, or large assemblages of cetaceans are observed near a vessel. All vessels must maintain a minimum separation distance of 100 m from sperm whales and all other baleen whales. All vessels must, to the maximum extent practicable, attempt to maintain a minimum separation distance of 50 m from all other marine mammals, with an understanding that at times this may not be possible (
e.g.,
for animals that approach the vessel).
When marine mammals are sighted while a vessel is underway, the vessel shall take action as necessary to avoid violating the relevant separation distance (
e.g.,
attempt to remain parallel to the animal's course, avoid excessive speed or abrupt changes in direction until the animal has left the area). If marine mammals are sighted within the relevant separation distance, the vessel must reduce speed and shift the engine to neutral, not engaging the engines until animals are clear of the area. This does not apply to any vessel towing gear or any vessel that is navigationally constrained.
Based on our evaluation of the applicant's proposed measures, NMFS has preliminarily determined that the proposed mitigation measures provide the means of effecting the least practicable impact on the affected species or stocks and their habitat, paying particular attention to rookeries, mating grounds, and areas of similar significance.
Proposed Monitoring and Reporting
In order to issue an IHA for an activity, section 101(a)(5)(D) of the MMPA states that NMFS must set forth requirements pertaining to the monitoring and reporting of such taking. The MMPA implementing regulations at 50 CFR 216.104(a)(13) indicate that requests for authorizations must include the suggested means of accomplishing the necessary monitoring and reporting that will result in increased knowledge of the species and of the level of taking or impacts on populations of marine mammals that are expected to be present while conducting the activities. Effective reporting is critical both to compliance as well as ensuring that the most value is obtained from the required monitoring.
Monitoring and reporting requirements prescribed by NMFS should contribute to improved understanding of one or more of the following:
Occurrence of marine mammal species or stocks in the area in which take is anticipated (e.g.,
presence, abundance, distribution, density);
Nature, scope, or context of likely marine mammal exposure to potential stressors/impacts (individual or cumulative, acute or chronic), through better understanding of: (1) action or environment (e.g.,
source characterization, propagation, ambient noise); (2) affected species (
e.g.,
life history, dive patterns); (3) co-occurrence of marine mammal species with the activity; or (4) biological or behavioral context of exposure (
e.g.,
age, calving or feeding areas);
Individual marine mammal responses (behavioral or physiological) to acoustic stressors (acute, chronic, or cumulative), other stressors, or cumulative impacts from multiple stressors;
How anticipated responses to stressors impact either: (1) long-term fitness and survival of individual marine mammals; or (2) populations, species, or stocks;
Effects on marine mammal habitat (e.g.,
marine mammal prey species, acoustic habitat, or other important physical components of marine mammal habitat); and,
Mitigation and monitoring effectiveness.
Vessel-Based Visual Monitoring
As described above, PSO observations would take place during daytime airgun operations. During seismic survey operations, at least five visual PSOs would be based aboard the
Langseth.
Two visual PSOs would be on duty at all times during daytime hours. The operator will work with the selected third-party observer provider to ensure PSOs have all equipment (including backup equipment) needed to adequately perform necessary tasks, including accurate determination of distance and bearing to observed marine mammals. L-DEO must use dedicated, trained, and NMFS-approved PSOs. At least one visual PSO aboard the vessel must have a minimum of 90 days at-sea experience working in those roles, respectively, with no more than 18 months elapsed since the conclusion of the at-sea experience. One visual PSO with such experience shall be designated as the lead for the entire protected species observation team. The lead PSO shall serve as primary point of contact for the vessel operator and ensure all PSO requirements per the IHA are met. To the maximum extent practicable, the experienced PSOs should be scheduled to be on duty with those PSOs with appropriate training but who have not yet gained relevant experience. The PSOs must have no tasks other than to conduct observational effort, record observational data, and communicate with and instruct relevant vessel crew with regard to the presence of marine mammals and mitigation requirements. PSO resumes shall be provided to NMFS for approval. Monitoring shall be conducted in accordance with the following requirements:
PSOs shall be independent, dedicated, trained visual PSOs and must be employed by a third-party observer provider;
PSOs shall have no tasks other than to conduct visual observational effort, collect data, and communicate with and instruct relevant vessel crew with regard to the presence of protected species and mitigation requirements (including brief alerts regarding maritime hazards); and
PSOs shall have successfully completed an approved PSO training
( printed page 19116)
course appropriate for their designated task (visual).
NMFS must review and approve PSO resumes accompanied by a relevant training course information packet that includes the name and qualifications (i.e.,
experience, training completed, or educational background) of the instructor(s), the course outline or syllabus, and course reference material as well as a document stating successful completion of the course;
PSOs must successfully complete relevant training, including completion of all required coursework and passing (80 percent or greater) a written and/or oral examination developed for the training program;
PSOs must have successfully attained a bachelor's degree from an accredited college or university with a major in one of the natural sciences, a minimum of 30 semester hours or equivalent in the biological sciences, and at least one undergraduate course in math or statistics; and
The educational requirements may be waived if the PSO has acquired the relevant skills through alternate experience. Requests for such a waiver shall be submitted to NMFS and must include written justification. Requests shall be granted or denied (with justification) by NMFS within 1 week of receipt of submitted information. Alternate experience that may be considered includes, but is not limited to (1) secondary education and/or experience comparable to PSO duties; (2) previous work experience conducting academic, commercial, or government-sponsored protected species surveys; or (3) previous work experience as a PSO; the PSO should demonstrate good standing and consistently good performance of PSO duties.
For data collection purposes, PSOs shall use standardized electronic data collection forms. PSOs shall record detailed information about any implementation of mitigation requirements, including the distance of animals to the airgun array and description of specific actions that ensued, the behavior of the animal(s), any observed changes in behavior before and after implementation of mitigation, and if shutdown was implemented, the length of time before any subsequent ramp-up of the airgun array. If required mitigation was not implemented, PSOs should record a description of the circumstances. At a minimum, the following information must be recorded:
○ Vessel name, vessel size and type, maximum speed capability of vessel;
○ Dates (MM/DD/YYYY) of departures and returns to port with port name;
○ PSO names and affiliations, PSO ID (initials or other identifier);
○ Date (MM/DD/YYYY) and participants of PSO briefings;
○ Visual monitoring equipment used (description);
○ PSO location on vessel and height (meters) of observation location above water surface;
○ Watch status (description);
○ Dates (MM/DD/YYYY) and times (Greenwich Mean Time (GMC)/Coordinated Universal Time (UTC)) of survey on/off effort and times (GMC/UTC) corresponding with PSO on/off effort;
○ Vessel location (decimal degrees) when survey effort began and ended and vessel location at beginning and end of visual PSO duty shifts;
○ Vessel location (decimal degrees) at 30-second intervals if obtainable from data collection software, otherwise at practical regular interval;
○ Vessel heading (compass heading) and speed (knots) at beginning and end of visual PSO duty shifts and upon any change;
○ Water depth (meters) (if obtainable from data collection software);
○ Environmental conditions while on visual survey (at beginning and end of PSO shift and whenever conditions changed significantly), including BSS and any other relevant weather conditions including cloud cover, fog, sun glare, and overall visibility to the horizon;
○ Factors that may have contributed to impaired observations during each PSO shift change or as needed as environmental conditions changed (description) (
e.g.,
vessel traffic, equipment malfunctions); and
○ Vessel/Survey activity information (and changes thereof) (description), such as airgun power output while in operation, number and volume of airguns operating in the array, tow depth of the array, and any other notes of significance (
i.e.,
pre-start clearance, ramp-up, shutdown, testing, shooting, ramp-up completion, end of operations, streamers,
etc.).
Upon visual observation of any marine mammals, the following information must be recorded:
○ Sighting ID (numeric);
○ Watch status (sighting made by PSO on/off effort, opportunistic, crew, alternate vessel/platform);
○ Location of PSO/observer (description);
○ Vessel activity at the time of the sighting (
e.g.,
deploying, recovering, testing, shooting, data acquisition, other);
○ PSO who sighted the animal/ID;
○ Time/date of sighting (GMT/UTC, MM/DD/YYYY);
○ Initial detection method (description);
○ Sighting cue (description);
○ Vessel location at time of sighting (decimal degrees);
○ Water depth (meters);
○ Direction of vessel's travel (compass direction);
○ Speed (knots) of the vessel from which the observation was made;
○ Direction of animal's travel relative to the vessel (description, compass heading);
○ Bearing to sighting (degrees);
○ Identification of the animal (
e.g.,
genus/species, lowest possible taxonomic level, or unidentified) and the composition of the group if there is a mix of species;
○ Species reliability (an indicator of confidence in identification) (1 = unsure/possible, 2 = probable, 3 = definite/sure, 9 = unknown/not recorded);
○ Estimated distance to the animal (meters) and method of estimating distance;
○ Estimated number of animals (high/low/best) (numeric);
○ Estimated number of animals by cohort (adults, yearlings, juveniles, calves, group composition,
etc.);
○ Description (as many distinguishing features as possible of each individual seen, including length, shape, color, pattern, scars or markings, shape and size of dorsal fin, shape of head, and blow characteristics);
○ Detailed behavior observations (
e.g.,
number of blows/breaths, number of surfaces, breaching, spyhopping, diving, feeding, traveling; as explicit and detailed as possible; note any observed changes in behavior);
○ Animal's closest point of approach (meters) and/or closest distance from any element of the airgun array;
○ Description of any actions implemented in response to the sighting (
e.g.,
delays, shutdown, ramp-up) and time and location of the action.
○ Photos (Yes/No);
○ Photo Frame Numbers (List of numbers); and
○ Conditions at time of sighting (Visibility; Beaufort Sea State).
Reporting
L-DEO shall submit a draft comprehensive report on all activities and monitoring results within 90 days of the completion of the survey or expiration of the IHA, whichever comes sooner. The report must describe all activities conducted and sightings of marine mammals, must provide full
( printed page 19117)
documentation of methods, results, and interpretation pertaining to all monitoring, and must summarize the dates and locations of survey operations and all marine mammal sightings (dates, times, locations, activities, associated survey activities). The draft report shall also include geo-referenced time-stamped vessel tracklines for all time periods during which airgun arrays were operating. Tracklines should include points recording any change in airgun array status (
e.g.,
when the sources began operating, when they were turned off, or when they changed operational status such as from full array to single gun or vice versa). Geographic Information System files shall be provided in Environmental Systems Research Institute shapefile format and include the UTC date and time, latitude in decimal degrees, and longitude in decimal degrees. All coordinates shall be referenced to the WGS84 geographic coordinate system. In addition to the report, all raw observational data shall be made available. The report must summarize data collected as described above in
Proposed Monitoring and Reporting.
A final report must be submitted within 30 days following resolution of any comments on the draft report.
Reporting Injured or Dead Marine Mammals
Discovery of injured or dead marine mammals
—In the event that personnel involved in the survey activities discover an injured or dead marine mammal, the L-DEO shall report the incident to the Office of Protected Resources (OPR) and NMFS as soon as feasible. The report must include the following information:
Time, date, and location (latitude/longitude) of the first discovery (and updated location information if known and applicable);
Species identification (if known) or description of the animal(s) involved;
Condition of the animal(s) (including carcass condition if the animal is dead);
Observed behaviors of the animal(s), if alive;
If available, photographs or video footage of the animal(s); and
General circumstances under which the animal was discovered.
Vessel strike
—In the event of a strike of a marine mammal by any vessel involved in the activities covered by the authorization, L-DEO shall report the incident to OPR and NMFS as soon as feasible. The report must include the following information:
Time, date, and location (latitude/longitude) of the incident;
Vessel's speed during and leading up to the incident;
Vessel's course/heading and what operations were being conducted (if applicable);
Status of all sound sources in use;
Description of avoidance measures/requirements that were in place at the time of the strike and what additional measure were taken, if any, to avoid strike;
Environmental conditions (e.g.,
wind speed and direction, BSS, cloud cover, visibility) immediately preceding the strike;
Species identification (if known) or description of the animal(s) involved;
Estimated size and length of the animal that was struck;
Description of the behavior of the marine mammal immediately preceding and following the strike;
If available, description of the presence and behavior of any other marine mammals present immediately preceding the strike;
Estimated fate of the animal (e.g.,
dead, injured but alive, injured and moving, blood or tissue observed in the water, status unknown, disappeared); and
To the extent practicable, photographs or video footage of the animal(s).
Negligible Impact Analysis and Determination
NMFS has defined negligible impact as an impact resulting from the specified activity that cannot be reasonably expected to, and is not reasonably likely to, adversely affect the species or stock through effects on annual rates of recruitment or survival (50 CFR 216.103). A negligible impact finding is based on the lack of likely adverse effects on annual rates of recruitment or survival (
i.e.,
population-level effects). An estimate of the number of takes alone is not enough information on which to base an impact determination. In addition to considering estimates of the number of marine mammals that might be “taken” through harassment, NMFS considers other factors, such as the likely nature of any impacts or responses (
e.g.,
intensity, duration), the context of any impacts or responses (
e.g.,
critical reproductive time or location, foraging impacts affecting energetics), as well as effects on habitat, and the likely effectiveness of the mitigation. We also assess the number, intensity, and context of estimated takes by evaluating this information relative to population status. Consistent with the 1989 preamble for NMFS' implementing regulations (54 FR 40338, September 29, 1989), the impacts from other past and ongoing anthropogenic activities are incorporated into this analysis via their impacts on the baseline (
e.g.,
as reflected in the regulatory status of the species, population size and growth rate where known, ongoing sources of human-caused mortality, or ambient noise levels).
To avoid repetition, the discussion of our analysis applies to all the species listed in table 1, given that the anticipated effects of this activity on these different marine mammal stocks are expected to be similar. NMFS does not anticipate that serious injury or mortality would occur as a result of L-DEO's planned survey, even in the absence of mitigation, and no serious injury or mortality is proposed to be authorized. As discussed in the
Potential Effects of Specified Activities on Marine Mammals and Their Habitat
section above, non-auditory physical effects and vessel strike are not expected to occur. NMFS expects that all potential take would be in the form of Level B behavioral harassment in the form of temporary avoidance of the area or decreased foraging (if such activity was occurring), responses that are considered to be of low severity, and with no lasting biological consequences (
e.g.,
Southall
et al.,
2007, 2021). These low-level impacts of behavioral harassment are not likely to impact the overall fitness of any individual or lead to population level effects of any species. As described above, Level A harassment is not expected to occur given the estimated small size of the Level A harassment zones.
In addition, the maximum expected Level B harassment zone around the survey vessel is 438 m. Therefore, the ensonified area surrounding the vessel is relatively small compared to the overall distribution of animals in the area and their use of the habitat. Feeding behavior is not likely to be significantly impacted as prey species are mobile and are broadly distributed throughout the survey area; therefore, marine mammals that may be temporarily displaced during survey activities are expected to be able to resume foraging once they have moved away from areas with disturbing levels of underwater noise. Because of the short duration (seven survey days) and temporary nature of the disturbance and the availability of similar habitat and resources in the surrounding area, the impacts to marine mammals and marine mammal prey species are not expected to cause significant or long-term fitness consequences for individual marine mammals or their populations.
( printed page 19118)
Additionally, the acoustic “footprint” of the proposed survey would be very small relative to the ranges of all marine mammals that would potentially be affected. Sound levels would increase in the marine environment in a relatively small area surrounding the vessel compared to the range of the marine mammals within the proposed survey area. The seismic array would be active 24 hours per day throughout the duration of the proposed survey. However, the very brief overall duration of the proposed survey (seven survey days) would further limit potential impacts that may occur as a result of the proposed activity.
Of the marine mammal species that are likely to occur in the project area, the following species are listed as endangered or threatened under the ESA: humpback whales (Central America DPS), fin whales, sei whales, sperm whales and Guadalupe fur seals. Additionally, the following species found in the action area that are not listed under the ESA but are depleted under the MMPA include: Pantropical spotted dolphin and spinner dolphin. The take numbers proposed for authorization for these species (table 6) are minimal relative to their modeled population sizes; therefore, we do not expect population-level impacts to any of these species. Moreover, the actual range of the populations extends past the area covered by the model, so modeled population sizes are likely smaller than their actual population size. Lastly, as previously described, meaningful impacts from the seismic survey are even less likely to occur for high-frequency cetaceans (
e.g.,
delphinids), as this group is relatively insensitive to sound produced at the predominant frequencies in an airgun pulse. Additionally for both high frequency and pinniped species the level A harassment isopleth is less than 5 m. The other marine mammal species that may be taken by harassment during L-DEO's seismic survey are not listed as threatened or endangered under the ESA or depleted under the MMPA. There is no designated critical habitat for any ESA-listed marine mammals within the project area.
There are no rookeries, mating, or calving grounds known to be biologically important to marine mammals within the survey area, and there are no feeding areas known to be biologically important to marine mammals within the survey area.
In summary and as described above, the following factors primarily support our preliminary determination that the impacts resulting from this activity are not expected to adversely affect any of the species or stocks through effects on annual rates of recruitment or survival:
No Level A harassment, serious injury, or mortality is anticipated or authorized;
The proposed activity is temporary and of relatively short duration (24 days total with 7 days of planned survey activity);
The anticipated impacts of the proposed activity on marine mammals would be temporary behavioral changes due to avoidance of the ensonified area, which is relatively small (see tables 4 and 5);
The availability of alternative areas of similar habitat value for marine mammals to temporarily vacate the survey area during the proposed survey to avoid exposure to sounds from the activity is readily abundant;
The potential adverse effects on fish or invertebrate species that serve as prey species for marine mammals from the proposed survey would be temporary and spatially limited and impacts to marine mammal foraging would be minimal; and,
The proposed mitigation measures are expected to reduce the number and severity of takes, to the extent practicable, by visually and/or acoustically detecting marine mammals within the established zones and implementing corresponding mitigation measures (e.g.,
delay; shutdown).
Based on the analysis contained herein of the likely effects of the specified activity on marine mammals and their habitat, and taking into consideration the implementation of the proposed monitoring and mitigation measures, NMFS preliminarily finds that the total marine mammal take from the proposed activity will have a negligible impact on all affected marine mammal species or stocks.
Small Numbers
As noted previously, only take of small numbers of marine mammals may be authorized under sections 101(a)(5)(A) and (D) of the MMPA for specified activities other than military readiness activities. The MMPA does not define small numbers and so, in practice, where estimated numbers are available, NMFS compares the number of individuals taken to the most appropriate estimation of abundance of the relevant species or stock in our determination of whether an authorization is limited to small numbers of marine mammals. When the predicted number of individuals to be taken is fewer than one-third of the species or stock abundance, the take is considered to be of small numbers. Additionally, other qualitative factors may be considered in the analysis, such as the temporal or spatial scale of the activities.
The number of takes NMFS proposes to authorize is below one-third of the modeled abundance for all relevant populations (specifically, take of individuals is less than 2 percent of the modeled abundance of each affected population, see table 6). This is conservative because the modeled abundance represents a population of the species and we assume all takes are of different individual animals, which is likely not the case. Some individuals may be encountered multiple times in a day, but PSOs would count them as separate individuals if they cannot be identified.
Based on the analysis contained herein of the proposed activity (including the proposed mitigation and monitoring measures) and the anticipated take of marine mammals, NMFS preliminarily finds that small numbers of marine mammals would be taken relative to the population size of the affected species or stocks.
Unmitigable Adverse Impact Analysis and Determination
There are no relevant subsistence uses of the affected marine mammal stocks or species implicated by this action. Therefore, NMFS has determined that the total taking of affected species or stocks would not have an unmitigable adverse impact on the availability of such species or stocks for taking for subsistence purposes.
Endangered Species Act
Section 7(a)(2) of the ESA of 1973 (16 U.S.C. 1531et seq.) requires that each Federal agency insure that any action it authorizes, funds, or carries out is not likely to jeopardize the continued existence of any endangered or threatened species or result in the destruction or adverse modification of designated critical habitat. To ensure ESA compliance for the issuance of IHAs, NMFS consults internally whenever we propose to authorize take for endangered or threatened species.
NMFS is proposing to authorize take of humpback whales (Central America DPS), fin whales, sei whales, sperm whales and Guadalupe fur seals, which are listed under the ESA. The NMFS OPR Permits and Conservation Division has requested initiation of section 7 consultation with the OPR ESA Interagency Cooperation Division for the issuance of this IHA. NMFS will conclude the ESA consultation prior to reaching a determination regarding the proposed issuance of the authorization.
( printed page 19119)
We request comment on our analyses, the proposed authorization, and any other aspect of this notice of proposed IHA for the proposed marine geophysical survey. We also request comment on the potential renewal of this proposed IHA as described in the paragraph below. Please include with your comments any supporting data or literature citations to help inform decisions on the request for this IHA or a subsequent renewal IHA.
On a case-by-case basis, NMFS may issue a one-time, 1-year renewal IHA following notice to the public providing an additional 15 days for public comments when (1) up to another year of identical or nearly identical activities as described in the
Description of Proposed Activity
section of this notice is planned or (2) the activities as described in the
Description of Proposed Activity
section of this notice would not be completed by the time the IHA expires and a renewal would allow for completion of the activities beyond that described in the
Dates and Duration
section of this notice, provided all of the following conditions are met:
A request for renewal is received no later than 60 days prior to the needed renewal IHA effective date (recognizing that the renewal IHA expiration date cannot extend beyond 1 year from expiration of the initial IHA).
The request for renewal must include the following:
(1) An explanation that the activities to be conducted under the requested renewal IHA are identical to the activities analyzed under the initial IHA, are a subset of the activities, or include changes so minor (
e.g.,
reduction in pile size) that the changes do not affect the previous analyses, mitigation and monitoring requirements, or take estimates (with the exception of reducing the type or amount of take).
(2) A preliminary monitoring report showing the results of the required monitoring to date and an explanation showing that the monitoring results do not indicate impacts of a scale or nature not previously analyzed or authorized.
Upon review of the request for renewal, the status of the affected species or stocks, and any other pertinent information, NMFS determines that there are no more than minor changes in the activities, the mitigation and monitoring measures will remain the same and appropriate, and the findings in the initial IHA remain valid.
Dated: April 28, 2025.
Catherine Marzin,
Acting Director, Office of Protected Resources, National Marine Fisheries Service.
Footnotes
1.
Please refer to the information given previously (
Description of Active Acoustic Sound Sources) regarding sound, characteristics of sound types, and metrics used in this document.
Use this for formal legal and research references to the published document.
90 FR 19090
Web Citation
Suggested Web Citation
Use this when citing the archival web version of the document.
“Takes of Marine Mammals Incidental to Specified Activities; Taking Marine Mammals Incidental to a Marine Geophysical Survey Off Western Mexico in the Eastern Tropical Pacific Ocean,” thefederalregister.org (May 5, 2025), https://thefederalregister.org/documents/2025-07613/takes-of-marine-mammals-incidental-to-specified-activities-taking-marine-mammals-incidental-to-a-marine-geophysical-surv.